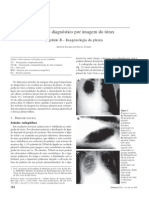
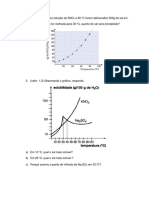
You might also like
- Guia de Dermatologia - SMS Campinas/SPDocument55 pagesGuia de Dermatologia - SMS Campinas/SPdmasetto100% (3)
- Atualização Das Diretrizes Brasileiras de Valvopatias - 2020Document56 pagesAtualização Das Diretrizes Brasileiras de Valvopatias - 2020João Marcos Lenhardt SilvaNo ratings yet
- Miotomos PrincipaisDocument1 pageMiotomos PrincipaisJoão Marcos Lenhardt SilvaNo ratings yet
- Guia de Dermatologia - SMS Campinas/SPDocument55 pagesGuia de Dermatologia - SMS Campinas/SPdmasetto100% (3)
- Lista Medicacao SUSDocument5 pagesLista Medicacao SUSKaio ViníciusNo ratings yet
- Lista Medicacao SUSDocument5 pagesLista Medicacao SUSKaio ViníciusNo ratings yet
- Atualização Das Diretrizes Brasileiras de Valvopatias - 2020Document56 pagesAtualização Das Diretrizes Brasileiras de Valvopatias - 2020João Marcos Lenhardt SilvaNo ratings yet
- Pancreatiteaguda2015 150728115931 Lva1 App6891 PDFDocument60 pagesPancreatiteaguda2015 150728115931 Lva1 App6891 PDFJoão Marcos Lenhardt SilvaNo ratings yet
- Hi Per SensiDocument29 pagesHi Per SensiMoura GomesNo ratings yet
- Bactérias Associadas Às Infecções Gastrointestinais II1Document48 pagesBactérias Associadas Às Infecções Gastrointestinais II1João Marcos Lenhardt SilvaNo ratings yet
- Seminário NeurologiaDocument171 pagesSeminário NeurologiaJoão Marcos Lenhardt SilvaNo ratings yet
- Caso Clnico - Sistema Respiratrio PDFDocument2 pagesCaso Clnico - Sistema Respiratrio PDFJoão Marcos Lenhardt SilvaNo ratings yet
- Cardiomiopatia PortifólioDocument16 pagesCardiomiopatia PortifólioJoão Marcos Lenhardt SilvaNo ratings yet
- Cinética Química - ManualDocument6 pagesCinética Química - ManualJoão Marcos Lenhardt SilvaNo ratings yet
- Resumo Histologia - Prova 1Document3 pagesResumo Histologia - Prova 1João Marcos Lenhardt SilvaNo ratings yet
- Caso ClinicoDocument2 pagesCaso ClinicoJoão Marcos Lenhardt SilvaNo ratings yet
- Funções Oxigenadas e NitrogenadasDocument6 pagesFunções Oxigenadas e NitrogenadasfreemanNo ratings yet
- Glossário de Termos Médicos para Acadêmicos de MedicinaDocument13 pagesGlossário de Termos Médicos para Acadêmicos de MedicinaJoão Marcos Lenhardt SilvaNo ratings yet
- Esquistossomose ResumoDocument4 pagesEsquistossomose ResumoJoão Marcos Lenhardt SilvaNo ratings yet
- Matematica Financeira No Enem Regra de TresDocument89 pagesMatematica Financeira No Enem Regra de TresÉrica TexNo ratings yet
- Texto 1 para 06.04 - Reinaldo Furlan - Uma Revisão Sobre A Filosofia Da CienciaDocument14 pagesTexto 1 para 06.04 - Reinaldo Furlan - Uma Revisão Sobre A Filosofia Da CienciaJoão Marcos Lenhardt SilvaNo ratings yet
- Atlas Digital de Histologia Basica PDFDocument75 pagesAtlas Digital de Histologia Basica PDFSabrina Baltazar100% (1)
- Abnt ProntoDocument29 pagesAbnt ProntoJoão Marcos Lenhardt SilvaNo ratings yet
- Cap 2. PleuraDocument12 pagesCap 2. PleuraEdvan CameloNo ratings yet
- Pilhas ExerciciosDocument37 pagesPilhas ExerciciosVinicius Garcia RibeiroNo ratings yet
- Coeficiente de Solubilidade - Exercicios - CursinhoDocument2 pagesCoeficiente de Solubilidade - Exercicios - CursinhoJoão Marcos Lenhardt SilvaNo ratings yet
- Terminologia Cirúrgica - DiegoDocument6 pagesTerminologia Cirúrgica - DiegoJoão Marcos Lenhardt SilvaNo ratings yet
- Modelo de Anamnese (Para Alunos) - Sandro - 1o Semestre THAM 2018.1 (T5)Document6 pagesModelo de Anamnese (Para Alunos) - Sandro - 1o Semestre THAM 2018.1 (T5)João Marcos Lenhardt SilvaNo ratings yet
- Radioatividade-Ensino Médio PDFDocument50 pagesRadioatividade-Ensino Médio PDFJoão Marcos Lenhardt SilvaNo ratings yet
- EXERCÍCIOS de Tabela Periódica - 20130319130048Document6 pagesEXERCÍCIOS de Tabela Periódica - 20130319130048Sonya KarlaNo ratings yet
- Como Usar A HP-50g para CalculoDocument12 pagesComo Usar A HP-50g para CalculoEliezerNo ratings yet
- Complexacao ESALQ PDFDocument176 pagesComplexacao ESALQ PDFGaby Salazar Mogollón100% (1)
- MATEMÁTICADocument118 pagesMATEMÁTICASuelen SantosNo ratings yet
- Revisao SimuladoDocument2 pagesRevisao SimuladovalimjosineiNo ratings yet
- Resumo - BiomecanicaDocument9 pagesResumo - BiomecanicaSandraMorethNo ratings yet
- Lista de Trabalhos 4 - CiclonagemDocument3 pagesLista de Trabalhos 4 - CiclonagemLuana Macedo100% (1)
- Relatorio de Produção Magnésio MetálicoDocument39 pagesRelatorio de Produção Magnésio MetálicoSidney OliveiraNo ratings yet
- Slides de Tecnologia Mecanica 17.07.18BDocument5 pagesSlides de Tecnologia Mecanica 17.07.18BAlfredo AlarNo ratings yet
- Aula 9Document40 pagesAula 9William Breternitz100% (1)
- Correção FP Bancos de CapacitoresDocument7 pagesCorreção FP Bancos de Capacitoreselton100% (1)
- 367891-Exercício PascalzimDocument4 pages367891-Exercício PascalzimDiego OliveiraNo ratings yet
- Artigo Edao-Comp - Reg - Primária - Cenários - Baixa - Hidraulicidade PDFDocument15 pagesArtigo Edao-Comp - Reg - Primária - Cenários - Baixa - Hidraulicidade PDFgdcerq8134No ratings yet
- Unidade 2 Desenvolvimento de Software para WebDocument8 pagesUnidade 2 Desenvolvimento de Software para WebGustavo DE PaulaNo ratings yet
- Stanadyne DE10 Pump (001-050) .En - PTDocument50 pagesStanadyne DE10 Pump (001-050) .En - PTwillyan100% (2)
- Manual Usuario - Manual DVDocument5 pagesManual Usuario - Manual DVGisseli MontanariNo ratings yet
- 3 - Estática Dos Fluidos - 2Document29 pages3 - Estática Dos Fluidos - 2Eliziane LeonNo ratings yet
- Ficha de Trabalho - Tabela Periódica - 10º Ano QuímicaDocument6 pagesFicha de Trabalho - Tabela Periódica - 10º Ano QuímicaTeresa CaldeiraNo ratings yet
- Destilação DiferencialDocument17 pagesDestilação DiferencialLuis Guilherme Foresto100% (1)
- Exercício 1 - Calculo NuméricoDocument4 pagesExercício 1 - Calculo Numéricotiago alexandreNo ratings yet
- PFD BPA Final-Páginas-1Document1 pagePFD BPA Final-Páginas-1Luiz Rodrigo AssisNo ratings yet
- Densimetria de Materiais GranuladoDocument5 pagesDensimetria de Materiais GranuladoGirlane SantosNo ratings yet
- Lista de Exercícios - SoluçõesDocument3 pagesLista de Exercícios - SoluçõesJeane Rodrigues BrederNo ratings yet
- Ats2 PDFDocument12 pagesAts2 PDFUenis Pinto TavaresNo ratings yet
- Oficina Pedagogica - PLANO DE AULADocument5 pagesOficina Pedagogica - PLANO DE AULAMara SilvaNo ratings yet
- Aula 1 - Introdução A Estradas de Rodagem e Projeto GeométricoDocument51 pagesAula 1 - Introdução A Estradas de Rodagem e Projeto GeométricoFabrício GuimarãesNo ratings yet
- 1 Análise Crítica Da Filosofia de WittgensteinDocument2 pages1 Análise Crítica Da Filosofia de WittgensteinRoberto BragaNo ratings yet
- Manual Sistema DGPark WEB Rev1 PDFDocument10 pagesManual Sistema DGPark WEB Rev1 PDFDomParking MinhaVagaNo ratings yet
- Apostila de Brigada de EmergênciaDocument37 pagesApostila de Brigada de EmergênciaLucas SerranoNo ratings yet
- Colaborar - Av1 - Linguagem Orientada A ObjetosDocument3 pagesColaborar - Av1 - Linguagem Orientada A Objetoselyasmin007No ratings yet
- Ap Matemática 6 Série EfDocument304 pagesAp Matemática 6 Série EfAlexandre MarianoNo ratings yet