You might also like
- AcuaporinasDocument40 pagesAcuaporinasAnonymous Mz32Sxwpcr100% (1)
- AcuaporinasDocument12 pagesAcuaporinasPeter P. R. MoralesNo ratings yet
- AcuaporinasDocument21 pagesAcuaporinasChristian Randy Castillo EnríquezNo ratings yet
- Informe 4-G7Document4 pagesInforme 4-G7Kattia Guizado AtaramaNo ratings yet
- Permeabilidad de La Membrana Celular Osmosis y Presión OsmóticaDocument7 pagesPermeabilidad de La Membrana Celular Osmosis y Presión OsmóticaFredy Ortiz100% (1)
- Semiología Médica Clase de Acidosis y Alcalosis MetabólicaDocument40 pagesSemiología Médica Clase de Acidosis y Alcalosis MetabólicaLuz Ledesma OropezaNo ratings yet
- Embriología Final 3Document220 pagesEmbriología Final 3Alexandra & ElvisNo ratings yet
- Cuadro Sinoptico de Filtracion Glomerular FisioSistemica. Sherlyn MatusDocument5 pagesCuadro Sinoptico de Filtracion Glomerular FisioSistemica. Sherlyn MatusSherlyn Matus Rios100% (1)
- Pancreas PDFDocument8 pagesPancreas PDFEdinson Cortez VasquezNo ratings yet
- Triada de Charcot y Pentada de ReynoldsDocument1 pageTriada de Charcot y Pentada de ReynoldsAgustin Bolaños SaltosNo ratings yet
- Capitulo 2-Composicion y Estructura de Las ProteinasDocument36 pagesCapitulo 2-Composicion y Estructura de Las Proteinasjuan sebastian parra boadaNo ratings yet
- Ensamblaje de Las ProteinasDocument29 pagesEnsamblaje de Las ProteinasAlejandro Mogrovejo50% (2)
- Hongo AureobasidiumDocument1 pageHongo AureobasidiumcristinaNo ratings yet
- Hidro CartografiaDocument3 pagesHidro CartografiaVictor Orihuela DíazNo ratings yet
- Ion Calcio Como Segundo MensajeroDocument19 pagesIon Calcio Como Segundo MensajeroIbarra martinez100% (19)
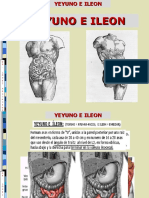
- Yeyuno e IleonDocument34 pagesYeyuno e IleonEdson Barba Patiño100% (2)
- IrrigacionDocument40 pagesIrrigacionYesica Quilca MamaniNo ratings yet
- Usmp Practica - Histologia 2018 - Tejido NerviosoDocument36 pagesUsmp Practica - Histologia 2018 - Tejido NerviosoJocelyn Aranda RabanalNo ratings yet
- Tejido EpitelialDocument71 pagesTejido EpitelialLuis Castañeda ZavaletaNo ratings yet
- Histología II Imágenes Primera Unidad 2016Document8 pagesHistología II Imágenes Primera Unidad 2016Christian Ballena RojasNo ratings yet
- Timo y Bazo ResultadosDocument2 pagesTimo y Bazo ResultadosJimmy Patricio Campoverde ErraizNo ratings yet
- Actividad de Las GrasasDocument4 pagesActividad de Las Grasasclaudia marcela rubio rojasNo ratings yet
- Pelvis EspinozaDocument131 pagesPelvis EspinozaRenzo RiveraNo ratings yet
- Guía Fase 2-1-18Document18 pagesGuía Fase 2-1-18Alejandra Alpaca RiverosNo ratings yet
- Inmunoglobulinas y Anticuerpos1Document13 pagesInmunoglobulinas y Anticuerpos1Luis Angel Ospina Herrera100% (1)
- Moleculas BiologicasDocument8 pagesMoleculas BiologicasWen BlancarteNo ratings yet
- Parcial 1 Bioqui 1Document2 pagesParcial 1 Bioqui 1Saul NicondNo ratings yet
- Práctica 11-Digestión de CarbohidrátosDocument12 pagesPráctica 11-Digestión de CarbohidrátosKEVIN JUNIOR RIVERA VASQUEZ100% (1)
- Nociones Basicas ElectrocardiogramaDocument4 pagesNociones Basicas ElectrocardiogramaAnayely RivasNo ratings yet
- Infome 7Document32 pagesInfome 7Victor Jesus Victorsg Siguas GarciaNo ratings yet
- Lab No1 Espectrofotometriax PDFDocument7 pagesLab No1 Espectrofotometriax PDFSolGimenezNo ratings yet
- La Extracción de ADN Se Realiza en Tres EtapasDocument3 pagesLa Extracción de ADN Se Realiza en Tres Etapassandra_muñoz_15100% (1)
- Clase 3 Filtracion Glomerular Medicina 2017Document28 pagesClase 3 Filtracion Glomerular Medicina 2017Antonia bugueñoNo ratings yet
- Glándulas Anexas de La PielDocument18 pagesGlándulas Anexas de La Pielrenato50% (2)
- Clase 03 ERITROPOYESIS PDFDocument34 pagesClase 03 ERITROPOYESIS PDFJuan Yarleque VilcaNo ratings yet
- Líquido TisularDocument44 pagesLíquido TisularBryant Manuel Ortiz Aquino100% (1)
- Los Aminoácidos y Las Proteínas Como Electrolitos y Su Capacidad Amortiguadora en El Organismo HumanoDocument12 pagesLos Aminoácidos y Las Proteínas Como Electrolitos y Su Capacidad Amortiguadora en El Organismo HumanoMiguel Ángel Jesús Lavarello BermejoNo ratings yet
- Celda VesicalDocument1 pageCelda VesicalvictoriaNo ratings yet
- Derivados Del Conducto MesonéfricoDocument2 pagesDerivados Del Conducto MesonéfricoLuccio RomaníNo ratings yet
- Monografia Ciclo Celular OficialDocument4 pagesMonografia Ciclo Celular OficialCv LiceliNo ratings yet
- Presentación 1Document15 pagesPresentación 1DianitaC.AvilaNo ratings yet
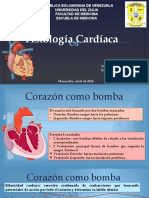
- Fisiologia CardíacaDocument76 pagesFisiologia Cardíacamade pernaNo ratings yet
- Fisiologia Renal 1Document80 pagesFisiologia Renal 1AlienNo ratings yet
- Control Local y Humoral Del Flujo Sanguíneo Por Los TejidosDocument46 pagesControl Local y Humoral Del Flujo Sanguíneo Por Los TejidosErnesto Ortiz100% (2)
- Histología Del RiñónDocument5 pagesHistología Del RiñónNutri RicNo ratings yet
- Final Embriologia UcsurDocument37 pagesFinal Embriologia UcsurAlejandro Treneman100% (1)
- Semiologia Renal Tema 2pdfDocument11 pagesSemiologia Renal Tema 2pdfJuanjo LeytónNo ratings yet
- Seminario Ciclo de KrebsDocument20 pagesSeminario Ciclo de KrebsJhonatan Rodriguez Moya0% (1)
- VÍAS URINARIAS DR - Nixor 2019Document13 pagesVÍAS URINARIAS DR - Nixor 2019Carol Jennifer Núñez Campos100% (1)
- Informe Hueso OccipitalDocument4 pagesInforme Hueso OccipitalDania BaculimaNo ratings yet
- Conducto EyaculadorDocument6 pagesConducto EyaculadorBesijess Cuevas CervantesNo ratings yet
- INFORMEDocument3 pagesINFORMEGustavo Herrera MontenegroNo ratings yet
- Cap.17 Sistema Digestivo Tubo Digestivo EXPO 4Document80 pagesCap.17 Sistema Digestivo Tubo Digestivo EXPO 4Juan Carlos Ledo Garcia100% (1)
- Silabo Fisiología 2015 IIDocument16 pagesSilabo Fisiología 2015 IILiz Gómez HuertaNo ratings yet
- Transporte y MezclaDocument46 pagesTransporte y MezclaLEIDY MENDIETANo ratings yet
- Metabolismo de Los Acidos GrasosDocument46 pagesMetabolismo de Los Acidos GrasosRoosvelt Cruz Córdova100% (2)
- Psicologia MedicaDocument40 pagesPsicologia MedicaTommy SalgueiroNo ratings yet
- Musculos InfrahioideosDocument60 pagesMusculos Infrahioideosloquito2012100% (1)
- Practica 8 PDFDocument7 pagesPractica 8 PDFLisbeth Bernal AlarcónNo ratings yet
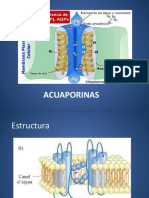
- Seminario 1 - AcuaporinasDocument8 pagesSeminario 1 - AcuaporinasDiego IgnacioNo ratings yet
- ANTAGONISMODocument6 pagesANTAGONISMOZoila ParedesNo ratings yet
- Bio SeguridadDocument10 pagesBio SeguridadZoila ParedesNo ratings yet
- 1 PB PDFDocument9 pages1 PB PDFNelson CarreraNo ratings yet
- Analisis GuatemalaDocument1 pageAnalisis GuatemalaZoila ParedesNo ratings yet
- Informe de FarmacologiaDocument2 pagesInforme de FarmacologiaZoila ParedesNo ratings yet
- ANTAGONISMODocument6 pagesANTAGONISMOZoila ParedesNo ratings yet
- Comunicación IntercelularDocument35 pagesComunicación IntercelularZoila ParedesNo ratings yet
- NeoplasiaDocument26 pagesNeoplasiaZoila ParedesNo ratings yet
- Las Diversas Iglesias Historicas: Rompieron Con La Unidad de La Iglesia de CristoDocument2 pagesLas Diversas Iglesias Historicas: Rompieron Con La Unidad de La Iglesia de CristoZoila ParedesNo ratings yet
- MusculoDocument2 pagesMusculoZoila ParedesNo ratings yet
- Inflamación y ReparaciónDocument3 pagesInflamación y ReparaciónZoila ParedesNo ratings yet
- Oído y Equilibrio 1Document20 pagesOído y Equilibrio 1Zoila ParedesNo ratings yet
- Aspectos Bioquimicos de La Caries DentalDocument18 pagesAspectos Bioquimicos de La Caries DentalZoila ParedesNo ratings yet
- Cadena Respiratoria IDocument39 pagesCadena Respiratoria IZoila ParedesNo ratings yet
- Criofractura y CriograbadoDocument12 pagesCriofractura y CriograbadoZoila Paredes0% (2)
- Clase Yesos Dentales PDFDocument54 pagesClase Yesos Dentales PDFZoila ParedesNo ratings yet
- Absorcion y Secrecion IntestinalDocument21 pagesAbsorcion y Secrecion IntestinalZoila ParedesNo ratings yet
- Aparato Cardiovascular DiaposDocument269 pagesAparato Cardiovascular DiaposZoila ParedesNo ratings yet
- Aadiapos CORRECCCIONDocument47 pagesAadiapos CORRECCCIONZoila ParedesNo ratings yet
- Acu A PorinasDocument30 pagesAcu A PorinasZoila ParedesNo ratings yet
- B-1 Bioquimica EnzimologiaDocument60 pagesB-1 Bioquimica EnzimologiaZoila ParedesNo ratings yet
- Acidos NucleicosDocument21 pagesAcidos NucleicosZoila ParedesNo ratings yet
- Mapa ConceptualDocument1 pageMapa ConceptualGabi CuevasNo ratings yet
- Fermentación - EcuRedDocument2 pagesFermentación - EcuRedjerryNo ratings yet
- Libro Completo ActinomicetosDocument251 pagesLibro Completo Actinomicetosichimatsu-sygunNo ratings yet
- Ingeniería GenéticaDocument8 pagesIngeniería Genéticav.valenzuela.baierNo ratings yet
- Factor Biótico Del Ecosistema para Primero de SecundariaDocument7 pagesFactor Biótico Del Ecosistema para Primero de SecundariaJoel FuentesNo ratings yet
- Ficha de Identificación de Trabajo de InvestigaciónDocument19 pagesFicha de Identificación de Trabajo de InvestigaciónROSINEIDE DE OLIVEIRA SILVANo ratings yet
- Cuadro Comparativo AtpDocument7 pagesCuadro Comparativo AtpKristian CobaNo ratings yet
- Valle Lucia R5 U3Document6 pagesValle Lucia R5 U3Luci Valle33% (6)
- El Protocolo Bredesen - Es La Nutrición La Clave Del AlzheimerDocument5 pagesEl Protocolo Bredesen - Es La Nutrición La Clave Del Alzheimerartcelis0% (1)
- Sesion N12 Ciencia y AmbienteDocument17 pagesSesion N12 Ciencia y AmbienteThaliacastilloguevarNo ratings yet
- Alteraciones GenéticasDocument9 pagesAlteraciones GenéticasDiegoNo ratings yet
- Guas Prcticas para El Uso de InsulinaDocument100 pagesGuas Prcticas para El Uso de InsulinaIngrid MoralesNo ratings yet
- Asis - PerúDocument44 pagesAsis - PerúRenzo Ezio Carrion VairoNo ratings yet
- CAE Biología 2021Document13 pagesCAE Biología 2021Jandro BurctlaNo ratings yet
- BIOLOGIA Mallas Snna AREA02Document29 pagesBIOLOGIA Mallas Snna AREA02Luis CaguaNo ratings yet
- GTA 1.ISem. 7°.21 PDFDocument5 pagesGTA 1.ISem. 7°.21 PDFAns NovNo ratings yet
- (Martes) Semana 5 Día 2 - 13-04-2021Document5 pages(Martes) Semana 5 Día 2 - 13-04-2021ItaloJRNo ratings yet
- El Huevo de GallinaDocument3 pagesEl Huevo de GallinajealegrialNo ratings yet
- Glicación No Enzimatica de Hemoglobina GlicosiladaDocument2 pagesGlicación No Enzimatica de Hemoglobina GlicosiladaAnita BarriaaNo ratings yet
- Enfermeades Del Sistema InmunologicoDocument5 pagesEnfermeades Del Sistema Inmunologicomaria guerra de gonzalezNo ratings yet
- Informacion Sobre Neurobiologia de La Memoria ExposicionDocument6 pagesInformacion Sobre Neurobiologia de La Memoria ExposicionTú Ayuda AcadémicaNo ratings yet
- Cuestionario. Aparato DigestivoDocument6 pagesCuestionario. Aparato Digestivodaniela garnica castroNo ratings yet
- La Acción de La Amilasa Sobre El AlmidónDocument4 pagesLa Acción de La Amilasa Sobre El AlmidónKAREN GONZLEZNo ratings yet
- Tipos de RadicalesDocument3 pagesTipos de RadicalesDaniel Barreto100% (1)
- Martes de PruebaDocument4 pagesMartes de Pruebayelka marianaNo ratings yet
- Plan 4 Biologiall 4 P 54Document11 pagesPlan 4 Biologiall 4 P 54Jose Pedro Martinez CalderonNo ratings yet
- Principales Enfermedades en MaízDocument3 pagesPrincipales Enfermedades en MaízorlandoNo ratings yet
- Ejercicios Ciclo Celular y Division CelularDocument21 pagesEjercicios Ciclo Celular y Division CelularLorraine FitzgeraldlNo ratings yet
- PRIORIZACIÓN DE CONTENIDOS Ciencias NaturalesDocument9 pagesPRIORIZACIÓN DE CONTENIDOS Ciencias NaturalesjanettNo ratings yet
- CRPDocument2 pagesCRPJuan Marcos Martinez Zevallos100% (1)