You might also like
- El Drama de Morir Viejo A Los 17Document3 pagesEl Drama de Morir Viejo A Los 17Orlando Alva GálvezNo ratings yet
- Cuando El Corazon Sufre Por Un... ReticuloDocument3 pagesCuando El Corazon Sufre Por Un... ReticuloOrlando Alva GálvezNo ratings yet
- Cuando Sudar Te MataDocument3 pagesCuando Sudar Te MataOrlando Alva GálvezNo ratings yet
- Adaptaciones y Exaptaciones PDFDocument65 pagesAdaptaciones y Exaptaciones PDFOrlando Alva GálvezNo ratings yet
- 00 CitogeneticaDocument3 pages00 CitogeneticaOrlando Alva GálvezNo ratings yet
- Aprendiendo El Uso de La FuerzaDocument3 pagesAprendiendo El Uso de La FuerzaOrlando Alva GálvezNo ratings yet
- Cannabis, Papas Fritas y ChocolateDocument4 pagesCannabis, Papas Fritas y ChocolateOrlando Alva GálvezNo ratings yet
- 01 El Concepto de GeneticaDocument3 pages01 El Concepto de GeneticaOrlando Alva GálvezNo ratings yet
- 1 Mitos de La CreaciónDocument6 pages1 Mitos de La CreaciónOrlando Alva GálvezNo ratings yet
- Evolucion Del Color de La Piel PDFDocument7 pagesEvolucion Del Color de La Piel PDFOrlando Alva GálvezNo ratings yet
- 04 Extensiones Del MendelismoDocument3 pages04 Extensiones Del MendelismoOrlando Alva GálvezNo ratings yet
- 02 Alteraciones CromosómicasDocument3 pages02 Alteraciones CromosómicasOrlando Alva GálvezNo ratings yet
- GIC 14 Linneo y La Clasificacion PDFDocument5 pagesGIC 14 Linneo y La Clasificacion PDFOrlando Alva GálvezNo ratings yet
- El Principe PicaroDocument26 pagesEl Principe PicaroOrlando Alva Gálvez100% (2)
- 03 Herencia MendelianaDocument3 pages03 Herencia MendelianaOrlando Alva GálvezNo ratings yet
- Bioquimica de Los BesosDocument2 pagesBioquimica de Los BesosOrlando Alva Gálvez100% (1)
- Recombinación Genética Durante La MeiosisDocument5 pagesRecombinación Genética Durante La MeiosisOrlando Alva Gálvez100% (1)
- CCMC 01Document28 pagesCCMC 01Maria Gloria Palomeque ContrerasNo ratings yet
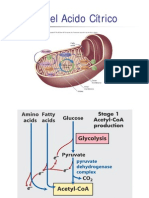
- Ciclo de KrebsDocument33 pagesCiclo de KrebsFrancisco SpitiaNo ratings yet
- Teoria de La Evolucion 2013Document19 pagesTeoria de La Evolucion 2013Jorge Tampan100% (1)
- Clase. Anomalias CromosomicasDocument107 pagesClase. Anomalias CromosomicasOrlando Alva GálvezNo ratings yet
- GIC 15 Darwin y La Evolucion PDFDocument5 pagesGIC 15 Darwin y La Evolucion PDFOrlando Alva GálvezNo ratings yet
- Bioquimica de AmorDocument2 pagesBioquimica de AmorOrlando Alva GálvezNo ratings yet
- Separata Cap, 1, 2 y 3 EnfoquesenterapiafamiliarsistémicaDocument16 pagesSeparata Cap, 1, 2 y 3 EnfoquesenterapiafamiliarsistémicaAntarel. abNo ratings yet
- 7.-TP y TTPaDocument11 pages7.-TP y TTPakorkotoNo ratings yet
- FILOSOFIA Entregable N°1 Carlos SuarezDocument9 pagesFILOSOFIA Entregable N°1 Carlos Suarezjuly jaimesNo ratings yet
- Matematica Financiera Dubraska PerezDocument2 pagesMatematica Financiera Dubraska Perezelpro25No ratings yet
- Modelo Educativo San MarcosDocument18 pagesModelo Educativo San MarcosAstrid Falla BarbozaNo ratings yet
- RESUMEN CAPITULO 2 - MarketingDocument11 pagesRESUMEN CAPITULO 2 - MarketingAndres AcevedoNo ratings yet
- Isaac Newton, físico y matemático inglésDocument6 pagesIsaac Newton, físico y matemático inglésKevin69No ratings yet
- Marleny Rincon - Psicoterapia No 2Document2 pagesMarleny Rincon - Psicoterapia No 2Siau San VicenteNo ratings yet
- Eje 2 - Seminario de InvestigacionDocument10 pagesEje 2 - Seminario de InvestigacionLessly Zaray MurilloNo ratings yet
- U0 - 03 - Prediccion y Comportamiento Del ReservorioDocument58 pagesU0 - 03 - Prediccion y Comportamiento Del ReservorioVictor MendozaNo ratings yet
- Medios de CultivoalumnosDocument11 pagesMedios de CultivoalumnosAnnie Martinez VazquezNo ratings yet
- Ejercicios Lenguaje de ProgramacionDocument7 pagesEjercicios Lenguaje de ProgramacionRené Suaña Coila50% (2)
- Preguntas Preparatorio Civil 2Document8 pagesPreguntas Preparatorio Civil 2Santiago Echavarria OlguinNo ratings yet
- Ejerccios MLSTRDocument12 pagesEjerccios MLSTRCarlosQuevedoNo ratings yet
- La Toponimia Como Fuente para El Estudio Del Territorio (2009)Document9 pagesLa Toponimia Como Fuente para El Estudio Del Territorio (2009)Pablo FolgueiraNo ratings yet
- Ronald DworkinDocument15 pagesRonald DworkinFlavio Eduardo Morales Reyes0% (1)
- Alicia Misrahi - Los Poderes de VenusDocument375 pagesAlicia Misrahi - Los Poderes de VenusMirtha Euridice Rodriguez UrbinaNo ratings yet
- Lectura Clase PMedio I NiemeyerDocument288 pagesLectura Clase PMedio I NiemeyerKaren RochaNo ratings yet
- Fundamentos Fisiologicos PDFDocument3 pagesFundamentos Fisiologicos PDFEddimar DiazNo ratings yet
- Ilceariadnaog Interaccionmedicamento Act1Document7 pagesIlceariadnaog Interaccionmedicamento Act1Ilse Ariadna Ordoñez GarciaNo ratings yet
- Manual de Inventario de Creencias IrracionalesDocument11 pagesManual de Inventario de Creencias IrracionalesPaola Mendoza Haurón Psicología - NeuropsicologíaNo ratings yet
- Clei 5Document27 pagesClei 5santi1212No ratings yet
- Protocolo Tecnico Cinetica PDFDocument14 pagesProtocolo Tecnico Cinetica PDFNana ChiquilloNo ratings yet
- Reporte de Tejupilco Contexto RuralDocument35 pagesReporte de Tejupilco Contexto RuralClaudia Gorethy Salinas AlmazanNo ratings yet
- Wesley, Spurgeon, Moody y el poder del EspírituDocument5 pagesWesley, Spurgeon, Moody y el poder del EspírituJosé Olviero MartínezNo ratings yet
- Lectura Marco Teorico Metodologico Variables OperacionalizacionDocument12 pagesLectura Marco Teorico Metodologico Variables OperacionalizacionJesús Dionisio Huarcaya JesúsNo ratings yet
- Fated To Be Alpha (Royal Rejection Book 1) - Skye AlderDocument98 pagesFated To Be Alpha (Royal Rejection Book 1) - Skye AlderRoberto Carlos OrellanaNo ratings yet
- Taller 1 EntropiaDocument7 pagesTaller 1 EntropiaGuillen Triana100% (2)
- Contrato GarnicaDocument3 pagesContrato GarnicaLeandro Jesus Arenas VeneroNo ratings yet
- Ses-Juev - Pl-Leemos Sobre El Día Nacional Del Ahorro de EnergíaDocument8 pagesSes-Juev - Pl-Leemos Sobre El Día Nacional Del Ahorro de EnergíaVictoriaPaulinaNo ratings yet