You might also like
- Manual Motor San Remo RecuperadoDocument68 pagesManual Motor San Remo RecuperadoDarlyn Iñiguez67% (6)
- Capacitacion de CianuroDocument36 pagesCapacitacion de CianuroLuis Christian Davila Cangalaya100% (2)
- Acuerdo 145 de 2008 Pensum Version 6Document7 pagesAcuerdo 145 de 2008 Pensum Version 6Facultad de Ingeniería Universidad de AntioquiaNo ratings yet
- Prospeccion Geologica de EsmeraldaDocument34 pagesProspeccion Geologica de EsmeraldaLusho SanchezNo ratings yet
- Laboratorio Lineas Equi-PotencialesDocument12 pagesLaboratorio Lineas Equi-PotencialesHarold.R GarciaNo ratings yet
- Resumen Guyton Capítulo 9Document5 pagesResumen Guyton Capítulo 9leslie ramirez80% (5)
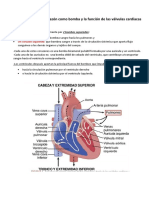
- Capítulo 9: Músculo Cardíaco: El Corazón Como Bomba y La Función de Las Válvulas CardíacasDocument37 pagesCapítulo 9: Músculo Cardíaco: El Corazón Como Bomba y La Función de Las Válvulas CardíacasManuela Monsalve TamarizNo ratings yet
- UABP 2. Músculo Cardíaco y Ciclo CardíacoDocument22 pagesUABP 2. Músculo Cardíaco y Ciclo CardíacoMartín PanicoNo ratings yet
- Informe Musculo LisoDocument21 pagesInforme Musculo LisoArturo TenorioNo ratings yet
- Cap 9 Músculo Cardíaco - El Corazón Como Bomba y La Función de Las Válvulas CardíacasDocument10 pagesCap 9 Músculo Cardíaco - El Corazón Como Bomba y La Función de Las Válvulas Cardíacasrodrigo4590No ratings yet
- Resumen Del Corazon VillacresDocument8 pagesResumen Del Corazon VillacresStalyn VillacresNo ratings yet
- Fisiologia Cardiaca GuytonDocument20 pagesFisiologia Cardiaca Guytonmate la femina100% (1)
- CAPITULO 9: Fisiologia MedicaDocument15 pagesCAPITULO 9: Fisiologia MedicaEsme MonroyNo ratings yet
- Fisiologia Del Sistema Cardiovascular Parte IDocument32 pagesFisiologia Del Sistema Cardiovascular Parte IJennifer Dayana Roa Umaña100% (1)
- Fisiología CardiacaDocument12 pagesFisiología CardiacaMiriam Loreley Cahuantzi FloresNo ratings yet
- Corazón Musculo Cardiaco ResumenesDocument16 pagesCorazón Musculo Cardiaco ResumenesLuciana BerriNo ratings yet
- Resumo Fisiologia IIDocument64 pagesResumo Fisiologia IIRebeca d'El-ReyNo ratings yet
- Apunte Fisio 2do Parcial 2018 PDFDocument103 pagesApunte Fisio 2do Parcial 2018 PDFIrina BaezNo ratings yet
- Fisiologia Del Musculo CardiacoDocument2 pagesFisiologia Del Musculo CardiacoPeña C. Angela R.No ratings yet
- Mecanismo Del CorazonDocument11 pagesMecanismo Del CorazonJc TovarNo ratings yet
- Potencial de Acción Prolongado en El Musculo Cardiaco: La Diferencia Del MusculoDocument6 pagesPotencial de Acción Prolongado en El Musculo Cardiaco: La Diferencia Del MusculoMaria Fernanda Carvajal CastillaNo ratings yet
- Actividad 5Document18 pagesActividad 5Suly VigoNo ratings yet
- Fisiologia Cardiaca GuytonDocument20 pagesFisiologia Cardiaca GuytonLuis LopezNo ratings yet
- Fisiología Del Sistema CirculatorioDocument20 pagesFisiología Del Sistema CirculatorioEmiliano Díaz CampanelliNo ratings yet
- Introduccion A La Fisiologia CardiacaDocument16 pagesIntroduccion A La Fisiologia CardiacaMartin PiattiNo ratings yet
- Musculo Cardiaco Preparatoria EvaDocument15 pagesMusculo Cardiaco Preparatoria Evanachocoolpro855No ratings yet
- Ciclo Cardíaco - MelgDocument7 pagesCiclo Cardíaco - MelgMELISA ELIZABETH LOPEZ GUERRERONo ratings yet
- Músculo CardíacoDocument15 pagesMúsculo CardíacoNara FlosanNo ratings yet
- 9 Músculo CardíacoDocument11 pages9 Músculo CardíacoServando Daniel Rivas BalderramaNo ratings yet
- Potencial de Accion Del Musculo Cardiaco y EkgDocument43 pagesPotencial de Accion Del Musculo Cardiaco y EkgPaola Custodio GonzalesNo ratings yet
- Propiedades Fisiológicas Y Estimulación Rítmica Del Corazón - Ok, WellDocument247 pagesPropiedades Fisiológicas Y Estimulación Rítmica Del Corazón - Ok, WellCarlos M.V.100% (3)
- El CorazónDocument11 pagesEl CorazónCaroline Menares CastilloNo ratings yet
- Está Formado Por Dos Bombas SeparadasDocument2 pagesEstá Formado Por Dos Bombas SeparadasAna JimenezNo ratings yet
- Acoplamiento Excitación-Contracioon Musculo CardíacoDocument2 pagesAcoplamiento Excitación-Contracioon Musculo CardíacoOriana BlancoNo ratings yet
- Electrocardiograma IIDocument9 pagesElectrocardiograma IIGeraldine LópezNo ratings yet
- Musculo Cardiaco Exposicion-1Document34 pagesMusculo Cardiaco Exposicion-1vickyNo ratings yet
- Conceptos Basicos en Fisiologia CardiovascularDocument26 pagesConceptos Basicos en Fisiologia CardiovascularBrenda Hernandez ValdezNo ratings yet
- Excitación y Contracción de Musculo Cardiaco PDFDocument5 pagesExcitación y Contracción de Musculo Cardiaco PDFCarolina FaleraNo ratings yet
- Ensayo FisiologiaDocument2 pagesEnsayo FisiologiaBelen FleitasNo ratings yet
- Arritmias CardiacasDocument16 pagesArritmias CardiacasJuan Ignacio GuerraNo ratings yet
- Cuestionario de Músculo Cardiaco (DIAZ VILCHIS ANDREA ERANDY)Document12 pagesCuestionario de Músculo Cardiaco (DIAZ VILCHIS ANDREA ERANDY)JR IbañezNo ratings yet
- Capitulo 9Document18 pagesCapitulo 9joelgomezbernardinoNo ratings yet
- Fisología - Músculo CardíacoDocument5 pagesFisología - Músculo CardíacoLucero Sonaly Mestanza LópezNo ratings yet
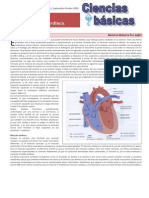
- Fisiología Cardiaca.: Revista Médica MD Número 3, Volumen 1 Septiembre-Octubre 2009Document4 pagesFisiología Cardiaca.: Revista Médica MD Número 3, Volumen 1 Septiembre-Octubre 2009soft220volNo ratings yet
- Corazón2 2 1Document47 pagesCorazón2 2 1Gabyta BarreraNo ratings yet
- Propiedades Del M.card. Del SapoDocument18 pagesPropiedades Del M.card. Del SapoGianfrancoNo ratings yet
- Potenciales de Acción en El Musculo CardiacoDocument5 pagesPotenciales de Acción en El Musculo Cardiacofelix GarcíaNo ratings yet
- CardiovascularDocument11 pagesCardiovascularWildriana RamosNo ratings yet
- Ilovepdf MergedDocument57 pagesIlovepdf MergedDavid Jesus Chacon KanterNo ratings yet
- Resumen CardioDocument1 pageResumen CardioKrystel AguayoNo ratings yet
- Resumo FisiologiaDocument11 pagesResumo FisiologiaRebeca d'El-ReyNo ratings yet
- Act.8.Resumen MusCard AbonceKarisa 301Document14 pagesAct.8.Resumen MusCard AbonceKarisa 301Andrea AbonceNo ratings yet
- Capitulo 9 GuytonDocument5 pagesCapitulo 9 Guytonbraham BarrosoNo ratings yet
- Fenomeno Mecanico Del CorazonDocument16 pagesFenomeno Mecanico Del Corazonkeiry Elibeth Peraza MajanoNo ratings yet
- Fisiologia CardiacaDocument7 pagesFisiologia CardiacaDonyell alexander Celis torresNo ratings yet
- Contracción MuscularDocument3 pagesContracción MuscularLillianDiazParedesNo ratings yet
- Musculo CardiacoDocument53 pagesMusculo CardiacoIdesRosangelaFerronatoOrtigoza67% (6)
- Cap 9 MUSCULO CARDIACODocument12 pagesCap 9 MUSCULO CARDIACOEsmeralda Pech AcostaNo ratings yet
- Capitulo 9 Resumen Guyton e Hall Tratado de Fisiologia MedicaDocument14 pagesCapitulo 9 Resumen Guyton e Hall Tratado de Fisiologia MedicaGilberto F. Fernandez100% (2)
- Cap 9 GuytonDocument11 pagesCap 9 GuytonAmbar Peña Mejia50% (2)
- Musculo CardíacoDocument9 pagesMusculo CardíacoTEIVA ALEJANDRO SALDANA FIGUEROANo ratings yet
- Introducción Básica a la Microcirculación de la Sangre: La Siguiente Generación en Cuidados de la SaludFrom EverandIntroducción Básica a la Microcirculación de la Sangre: La Siguiente Generación en Cuidados de la SaludNo ratings yet
- Piensa Antes De Beber: Las consecuencias de consumir bebidas alcohólicasFrom EverandPiensa Antes De Beber: Las consecuencias de consumir bebidas alcohólicasRating: 5 out of 5 stars5/5 (1)
- Mejorar la Circulación Sanguínea de Forma Natural: Sanar el Sistema Circulatorio y inmunológico para Prevenir Enfermedades, Aumentar la Energía, Aliviar el Estrés y Sentirte Mejor que NuncaFrom EverandMejorar la Circulación Sanguínea de Forma Natural: Sanar el Sistema Circulatorio y inmunológico para Prevenir Enfermedades, Aumentar la Energía, Aliviar el Estrés y Sentirte Mejor que NuncaRating: 4.5 out of 5 stars4.5/5 (7)
- Lab Oratorio Rocas 2016Document2 pagesLab Oratorio Rocas 2016Jose Luis Nava HernandezNo ratings yet
- Campo Magnético de Un Toroide y Solenoide Taller - LeccionDocument3 pagesCampo Magnético de Un Toroide y Solenoide Taller - Leccionwilly solisNo ratings yet
- EXAMEN 1RO BimestralDocument2 pagesEXAMEN 1RO BimestralMisael Zanabria HuanascaNo ratings yet
- Soldadura de AluminioDocument18 pagesSoldadura de Aluminioapi-118986524No ratings yet
- Lab - 1 Masa ResorteDocument3 pagesLab - 1 Masa Resortegarfacio30No ratings yet
- PC5500-D 15045 Operation and Maintenance Manual Spanish PDFDocument526 pagesPC5500-D 15045 Operation and Maintenance Manual Spanish PDFLuisAlbertoVerdejoTapia100% (1)
- SEMANA N01-2019-IiDocument4 pagesSEMANA N01-2019-IiBilly Deybi Niño MoralesNo ratings yet
- Informe Laboratorio Filtros ActivosDocument5 pagesInforme Laboratorio Filtros Activoscaromero270No ratings yet
- FisicaDocument3 pagesFisicaAlejandra M. Avila0% (1)
- Analogicos 1Document11 pagesAnalogicos 1Roberto Olmos PimentelNo ratings yet
- Introduccion de DestilacionDocument82 pagesIntroduccion de DestilacionOH SoeNo ratings yet
- F BS2 01 MatricesDocument16 pagesF BS2 01 MatricesLuciia Méndez GutiérrezNo ratings yet
- Calculo de Asentamientos en SuelosDocument12 pagesCalculo de Asentamientos en SuelosRoyerArias100% (5)
- Esfera de AlmacenamientoDocument18 pagesEsfera de AlmacenamientoEstuardo Javier Gan RodríguezNo ratings yet
- Propiedades MagneticasDocument11 pagesPropiedades MagneticasChristian GerardoNo ratings yet
- Integrales de SuperficieDocument14 pagesIntegrales de SuperficieuleciaNo ratings yet
- Teoría Atomica Laura MangaDocument18 pagesTeoría Atomica Laura MangaLaura MangaNo ratings yet
- Madera LaminadaDocument3 pagesMadera LaminadaSuarez F Slayper JhonsNo ratings yet
- Mas Problemas de Trabajo y Energia RDocument8 pagesMas Problemas de Trabajo y Energia RyarliNo ratings yet
- Itemizado IluminaciónDocument7 pagesItemizado IluminaciónAnonymous 1tmhviNo ratings yet
- IF4 AltaDocument9 pagesIF4 AltaAnthony Hbk JerichoNo ratings yet
- Estudio de Suelos Real Felipe (Q 1.97)Document16 pagesEstudio de Suelos Real Felipe (Q 1.97)pyxis_mlNo ratings yet
- Guias de Laboratorio Quimica BasicaDocument81 pagesGuias de Laboratorio Quimica BasicaAngel Remache100% (1)
- Ingenieria Biomedica (Tesis de Maestria)Document144 pagesIngenieria Biomedica (Tesis de Maestria)Vladimir Morote100% (2)
- AlcoholesDocument15 pagesAlcoholesFajardo Pinchi JanhitaNo ratings yet