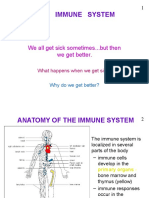
You might also like
- Visualization of Neutrophil Extra Cellular Traps in TEMDocument6 pagesVisualization of Neutrophil Extra Cellular Traps in TEMjose_gladiusNo ratings yet
- The Role of Histone Deacetylases in Asthma and Allergic Disease.Document5 pagesThe Role of Histone Deacetylases in Asthma and Allergic Disease.jose_gladiusNo ratings yet
- Singlet Oxygen Is Essential For Neutrophil Extra Cellular Trap FormationDocument5 pagesSinglet Oxygen Is Essential For Neutrophil Extra Cellular Trap Formationjose_gladiusNo ratings yet
- Potential Role of Epithelial Cell-Derived Histone H1 Proteins in Innate Antimicrobial Defense in The Human Gastrointestinal TractDocument10 pagesPotential Role of Epithelial Cell-Derived Histone H1 Proteins in Innate Antimicrobial Defense in The Human Gastrointestinal Tractjose_gladiusNo ratings yet
- The Proteomic Profile of Circulating Pentraxin 3 (PTX3) Complex in Sepsis Demonstrates The Interaction With Azurocidin 1 and Other Components of Neutrophil Extra Cellular TrapsDocument35 pagesThe Proteomic Profile of Circulating Pentraxin 3 (PTX3) Complex in Sepsis Demonstrates The Interaction With Azurocidin 1 and Other Components of Neutrophil Extra Cellular Trapsjose_gladiusNo ratings yet
- Non Type Able Haemophilus Influenzae Initiates Formation of Neutrophil Extra Cellular TrapsDocument8 pagesNon Type Able Haemophilus Influenzae Initiates Formation of Neutrophil Extra Cellular Trapsjose_gladiusNo ratings yet
- Specific Post-Translational Histone Modifications of Neutrophil Extra Cellular Traps As Immunogens and Potential Targets of Lupus Auto AntibodiesDocument14 pagesSpecific Post-Translational Histone Modifications of Neutrophil Extra Cellular Traps As Immunogens and Potential Targets of Lupus Auto Antibodiesjose_gladiusNo ratings yet
- Restoration of Anti-Aspergillus Defense by Neutrophil Extra Cellular Traps in Human Chronic Granulomatous Disease After Gene Therapy Is Calprotectin-DependentDocument17 pagesRestoration of Anti-Aspergillus Defense by Neutrophil Extra Cellular Traps in Human Chronic Granulomatous Disease After Gene Therapy Is Calprotectin-Dependentjose_gladiusNo ratings yet
- PAD4-Mediated Neutrophil Extra Cellular Trap Formation Is Not Required For Immunity Against Influenza InfectionDocument10 pagesPAD4-Mediated Neutrophil Extra Cellular Trap Formation Is Not Required For Immunity Against Influenza Infectionjose_gladiusNo ratings yet
- Neutrophil Extra Cellular Traps Directly Induce Epithelial and Endothelial Cell Death A Predominant Role of HistonesDocument14 pagesNeutrophil Extra Cellular Traps Directly Induce Epithelial and Endothelial Cell Death A Predominant Role of Histonesjose_gladiusNo ratings yet
- Neutrophil Extra Cellular Traps Capture and Kill Candida Albicans Yeast and Hyphal FormsDocument9 pagesNeutrophil Extra Cellular Traps Capture and Kill Candida Albicans Yeast and Hyphal Formsjose_gladiusNo ratings yet
- Neutrophil Extra Cellular Traps Casting The NET Over Pa Tho GenesisDocument5 pagesNeutrophil Extra Cellular Traps Casting The NET Over Pa Tho Genesisjose_gladiusNo ratings yet
- Molecular Chaperones and Cell Signalling - Henderson 2005Document345 pagesMolecular Chaperones and Cell Signalling - Henderson 2005jose_gladiusNo ratings yet
- Histone Deacetylases Transcriptional Control, and Cancer.Document16 pagesHistone Deacetylases Transcriptional Control, and Cancer.jose_gladiusNo ratings yet
- Environmental Epigenetics of Asthma and UpdateDocument13 pagesEnvironmental Epigenetics of Asthma and Updatejose_gladiusNo ratings yet
- Habitos de SeñalizacionDocument15 pagesHabitos de Señalizacionjose_gladiusNo ratings yet
- Environmental Epigenetics and AsthmaDocument7 pagesEnvironmental Epigenetics and Asthmajose_gladiusNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Actividad 2 Artículo 1Document10 pagesActividad 2 Artículo 1byronNo ratings yet
- Salmonella Infections Clinical Immunological and Molecular Aspects Advances in Molecular and Cellular Microbiology PDFDocument402 pagesSalmonella Infections Clinical Immunological and Molecular Aspects Advances in Molecular and Cellular Microbiology PDFFredNo ratings yet
- Fish ImmunologyDocument32 pagesFish ImmunologyRokhmadRofiqNo ratings yet
- National Dental College and Hospital: Seminar On ImmunityDocument24 pagesNational Dental College and Hospital: Seminar On Immunitydr parveen bathlaNo ratings yet
- Immunologic Mechanisms in AsthmaDocument9 pagesImmunologic Mechanisms in AsthmaKessi VikaneswariNo ratings yet
- Sleep Health Reciprocal Regulation of Sleep and Innate ImmunityDocument27 pagesSleep Health Reciprocal Regulation of Sleep and Innate ImmunityPolly MollyNo ratings yet
- Innate Immunity: - Properties of Innate Immunity - Components of Innate ImmunityDocument39 pagesInnate Immunity: - Properties of Innate Immunity - Components of Innate Immunityrima melliaNo ratings yet
- The Evolution of ChiropracticDocument122 pagesThe Evolution of ChiropracticsololuiNo ratings yet
- The Structure and Function of White Blood CellsDocument8 pagesThe Structure and Function of White Blood CellsNaiomiNo ratings yet
- 02 - Innate ImmunityDocument63 pages02 - Innate ImmunityUmar UsmanNo ratings yet
- CROHNSDocument2 pagesCROHNSAlvin Germo Pasuquin100% (1)
- Hoffman - Hematology. Basic Principles and Practice (3th Edition)Document117 pagesHoffman - Hematology. Basic Principles and Practice (3th Edition)Carmina Lepa100% (1)
- Immunity (Lecture Notes)Document5 pagesImmunity (Lecture Notes)Anisha NishanthNo ratings yet
- Yi Jin JingDocument17 pagesYi Jin Jingflorinhangan50% (2)
- Innate ImmunityDocument29 pagesInnate ImmunityAsad MirzaNo ratings yet
- Pathophysiology Test Bank CH 6-9Document33 pagesPathophysiology Test Bank CH 6-9Joyy86% (7)
- Infection and Host ResistanceDocument62 pagesInfection and Host ResistanceAndrea Marie T. Artajo-BoheromNo ratings yet
- ImmunityDocument44 pagesImmunityHam Za100% (1)
- Chapter Two Micro 303-محولDocument12 pagesChapter Two Micro 303-محولshimaa dewedarNo ratings yet
- Components of The Immune SystemDocument8 pagesComponents of The Immune Systemبراءة أحمد السلاماتNo ratings yet
- Immunobiology: 1 Week To Check With Me. Follow The Syllabus's Guidelines!!Document9 pagesImmunobiology: 1 Week To Check With Me. Follow The Syllabus's Guidelines!!Kay YgNo ratings yet
- Vitamin C and Viral InfectionDocument3 pagesVitamin C and Viral InfectionFARCASANU MARIA-ANDREEANo ratings yet
- BSI Natures Self DefenceDocument12 pagesBSI Natures Self DefencesreenitthiNo ratings yet
- ImmunologyDocument1 pageImmunologyDr.2020No ratings yet
- Project ImmunityDocument14 pagesProject ImmunityAshit VermaNo ratings yet
- Immuno Notes LectureDocument30 pagesImmuno Notes LectureSteph TabasaNo ratings yet
- Updated Ebook 'The Ultimate Guide To Preventing Acne'Document15 pagesUpdated Ebook 'The Ultimate Guide To Preventing Acne'c2fnascimentoNo ratings yet
- Encyclopedia of Medical Immunology - Autoimmune Diseases GoonerDocument1,327 pagesEncyclopedia of Medical Immunology - Autoimmune Diseases GoonerAleNo ratings yet
- Can Oral Sex Transmit HIV?Document52 pagesCan Oral Sex Transmit HIV?Leo LamsNo ratings yet
- L.Shevyrdyaeva English For Biology Workbook 1Document11 pagesL.Shevyrdyaeva English For Biology Workbook 1Магистры Биологи 2021-2023No ratings yet