You might also like
- Activity of Selected Aromatic Amino Acids in Biological SystemsDocument6 pagesActivity of Selected Aromatic Amino Acids in Biological SystemsEduardo Lopez MedranoNo ratings yet
- 14 CatecholamineDocument5 pages14 CatecholamineZiedTrikiNo ratings yet
- Resin UptakeDocument4 pagesResin UptakeCome BackNo ratings yet
- Phenylalanine and Tyrosine Metabolism (18 Oct)Document14 pagesPhenylalanine and Tyrosine Metabolism (18 Oct)api-19824406100% (6)
- Adrenergic Agents: Shengquan LiuDocument36 pagesAdrenergic Agents: Shengquan LiuAnaliza Kitongan LantayanNo ratings yet
- Antipsychotics 1Document25 pagesAntipsychotics 1bcloverasta905No ratings yet
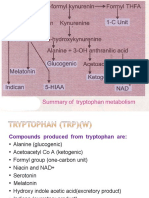
- Metabolism of TryptophanDocument38 pagesMetabolism of Tryptophanjagan mohan rao vanaNo ratings yet
- Running Head: Biochemistry Assignment 1Document6 pagesRunning Head: Biochemistry Assignment 1Fredrick LiyengaNo ratings yet
- Adrenaline PresentationDocument12 pagesAdrenaline PresentationAshley MudzingwaNo ratings yet
- Gene RegulationDocument86 pagesGene RegulationMeenal MeshramNo ratings yet
- Biochemistry of IsmDocument15 pagesBiochemistry of IsmHamed KamelNo ratings yet
- Answers To Case 38: Phenylketonuria (Pku) : Summary: A 1-Year-Old Girl Born Outside The United States With DevelopmentalDocument9 pagesAnswers To Case 38: Phenylketonuria (Pku) : Summary: A 1-Year-Old Girl Born Outside The United States With DevelopmentalKarilNo ratings yet
- Nucleotide Metabolism PPDocument65 pagesNucleotide Metabolism PPCLEMENTNo ratings yet
- Study Guide T2 BioChem2Document12 pagesStudy Guide T2 BioChem2janeNo ratings yet
- Dopa RewardDocument8 pagesDopa RewardDenia Méndez ÁlvarezNo ratings yet
- Purine Metabolism PDFDocument29 pagesPurine Metabolism PDFtrinitysugumar0% (1)
- Faktor Neurobiologi: Qara Syifa Fachrani 1610211039Document22 pagesFaktor Neurobiologi: Qara Syifa Fachrani 1610211039Qara Syifa FNo ratings yet
- Metabolism of Nucleoproteins Part IDocument50 pagesMetabolism of Nucleoproteins Part IAgafioNo ratings yet
- Biochemical Basis of Phenylketonuria and Maple Syrup SyndromeDocument5 pagesBiochemical Basis of Phenylketonuria and Maple Syrup SyndromeLauraTomicNo ratings yet
- PhenylalanineDocument4 pagesPhenylalaninemaithili hedaooNo ratings yet
- The Role and ApplicationDocument12 pagesThe Role and ApplicationÂn NguyễnNo ratings yet
- Neurophysiology of Depression and Bipolar Affective DisorderDocument17 pagesNeurophysiology of Depression and Bipolar Affective DisorderShivan A.C.No ratings yet
- Presentacion EntenderDocument37 pagesPresentacion EntenderJuan David Marin ChiguachiNo ratings yet
- Catecholamine: From Wikipedia, The Free EncyclopediaDocument6 pagesCatecholamine: From Wikipedia, The Free EncyclopediaHasrul MuslihNo ratings yet
- Dopamine, Serotonine, Epinephrine, and NorephinephrineDocument28 pagesDopamine, Serotonine, Epinephrine, and Norephinephrinehafshah100% (1)
- Neuro Nutrition 1983 v12n01 p038Document6 pagesNeuro Nutrition 1983 v12n01 p038LieMinkNo ratings yet
- Pharmacology of AutacoidsDocument13 pagesPharmacology of AutacoidsInocenteNo ratings yet
- Bylund 2014Document3 pagesBylund 2014Javier RiveraNo ratings yet
- Hormone Replacement in Men PDFDocument1 pageHormone Replacement in Men PDFPanser CueNo ratings yet
- New Vishani Tharindi 19041425graduation Thesis 20230522Document28 pagesNew Vishani Tharindi 19041425graduation Thesis 20230522vishpunchihewaNo ratings yet
- Phenylketonuria PathogenesisDocument2 pagesPhenylketonuria PathogenesisIrene CerisseNo ratings yet
- Egyptian CohortDocument16 pagesEgyptian Cohortesh.scicoNo ratings yet
- Trans of Harper's Illustrated Biochemistry CH 32Document3 pagesTrans of Harper's Illustrated Biochemistry CH 32Richelle Dianne Ramos-Giang100% (1)
- Influence of Antidepressant Drugs On EctDocument7 pagesInfluence of Antidepressant Drugs On EctconstanceNo ratings yet
- Catecholamine: Epinephrine (Adrenaline)Document7 pagesCatecholamine: Epinephrine (Adrenaline)Unggul YudhaNo ratings yet
- Ytochrome Nzymes: Presented by Deshmukh MD Faizan M. Pharm (1 Sem)Document20 pagesYtochrome Nzymes: Presented by Deshmukh MD Faizan M. Pharm (1 Sem)Anitha Mary DambaleNo ratings yet
- 10 Diversity of Endocrine System IIDocument23 pages10 Diversity of Endocrine System IIcutethoybaNo ratings yet
- Bio Chem Case ThreeDocument2 pagesBio Chem Case ThreeLeonard Rey Suratos ManabatNo ratings yet
- ScieDocument3 pagesSciellubitpauloNo ratings yet
- Mammalian Sperm MotilityDocument9 pagesMammalian Sperm MotilityLevvis AicragNo ratings yet
- Metabolism of TyrosineDocument1 pageMetabolism of TyrosineSophie Mwaniki NyihaNo ratings yet
- Farmakologi ParkinsonsDocument12 pagesFarmakologi ParkinsonsAnisa DwiiNo ratings yet
- DR Okunowo Wahab Introductory Molecular Biology Lecture Note I (Nucleotides Metabolism)Document20 pagesDR Okunowo Wahab Introductory Molecular Biology Lecture Note I (Nucleotides Metabolism)modelprof100% (2)
- Phosphorylation: DNA MethylationDocument2 pagesPhosphorylation: DNA MethylationCa Thy HuỳnhNo ratings yet
- BSC Sem VI Nuc Aid MetabDocument24 pagesBSC Sem VI Nuc Aid Metabstarboigaming08No ratings yet
- Case3 PhenylketunuriaDocument2 pagesCase3 PhenylketunuriaKath BoniolNo ratings yet
- TryptophanDocument41 pagesTryptophanmahalakshmiNo ratings yet
- Presentation of Proteomics and GenomicsDocument37 pagesPresentation of Proteomics and GenomicsJunaid MughalNo ratings yet
- Metabolism of NucleotidesDocument24 pagesMetabolism of NucleotidesAditya NayakNo ratings yet
- Reisser2002 Ic50Document4 pagesReisser2002 Ic50Jesús Rafael Méndez NateraNo ratings yet
- Phylogenetic Analysis of Phenylalanine Hydroxylase Enzyme and Its Future Aspect in Treatment of Phenylalanine Hydroxylase Enzyme Deficiency (Phenylketonuria)Document4 pagesPhylogenetic Analysis of Phenylalanine Hydroxylase Enzyme and Its Future Aspect in Treatment of Phenylalanine Hydroxylase Enzyme Deficiency (Phenylketonuria)International Journal of Innovative Science and Research TechnologyNo ratings yet
- Name: Jegathiswary A/P Ganasan Matrics: S56094 Code: BIO4102 Date: 11/07/2021Document2 pagesName: Jegathiswary A/P Ganasan Matrics: S56094 Code: BIO4102 Date: 11/07/2021Jegathiswary GanasanNo ratings yet
- s56094 Bio4102 Section BDocument2 pagess56094 Bio4102 Section BJegathiswary GanasanNo ratings yet
- AxolightDocument3 pagesAxolightnandaajiNo ratings yet
- FluoxetineDocument2 pagesFluoxetineMagdalena ArdeleanNo ratings yet
- Met Purin 28 OktDocument62 pagesMet Purin 28 OktXIID 67No ratings yet
- Wisner Et Al 2006Document6 pagesWisner Et Al 2006Lyka MahrNo ratings yet
- 19 SYNTHESIS OF ADRENALINEADRENERGIC RECEPTORS by COL AZMAT ALIDocument53 pages19 SYNTHESIS OF ADRENALINEADRENERGIC RECEPTORS by COL AZMAT ALIAhmed YTNo ratings yet
- Dopamin: Nur Fadhillah Khalid (P062181024) Mata Kuliah: NeurontrasmitterDocument27 pagesDopamin: Nur Fadhillah Khalid (P062181024) Mata Kuliah: Neurontrasmitternur fadhillah khalidNo ratings yet
- Catecholamine Research in the 21st Century: Abstracts and Graphical Abstracts, 10th International Catecholamine Symposium, 2012From EverandCatecholamine Research in the 21st Century: Abstracts and Graphical Abstracts, 10th International Catecholamine Symposium, 2012Lee E. EidenNo ratings yet
- Amanda N. Carey Et Al - Endogenous Kappa Opioid Activation Mediates Stress-Induced Deficits in Learning and MemoryDocument8 pagesAmanda N. Carey Et Al - Endogenous Kappa Opioid Activation Mediates Stress-Induced Deficits in Learning and MemoryNeerFamNo ratings yet
- Paul Aron - Hallucinogens and SchizophreniaDocument10 pagesPaul Aron - Hallucinogens and SchizophreniaNeerFamNo ratings yet
- Douglas J. Sheffler and Bryan L. Roth - Salvinorin A: The Magic Mint' Hallucinogen Finds A Molecular Target in The Kappa Opioid ReceptorDocument3 pagesDouglas J. Sheffler and Bryan L. Roth - Salvinorin A: The Magic Mint' Hallucinogen Finds A Molecular Target in The Kappa Opioid ReceptorNeerFamNo ratings yet
- Bryan L. Roth Et Al - Salvinorin A: A Potent Naturally Occurring Nonnitrogenous Kappa Opioid Selective AgonistDocument6 pagesBryan L. Roth Et Al - Salvinorin A: A Potent Naturally Occurring Nonnitrogenous Kappa Opioid Selective AgonistNeerFamNo ratings yet
- Deepak Cyril D'Souza Et Al-The Psychotomimetic Effects of Intravenous Delta-9-Tetrahydrocannabinol in Healthy Individuals: Implications For PsychosisDocument15 pagesDeepak Cyril D'Souza Et Al-The Psychotomimetic Effects of Intravenous Delta-9-Tetrahydrocannabinol in Healthy Individuals: Implications For PsychosisPoloGreenNo ratings yet
- Kevin Tidgewell Et Al - Synthesis of Salvinorin A Analogues As Opioid Receptor ProbesDocument5 pagesKevin Tidgewell Et Al - Synthesis of Salvinorin A Analogues As Opioid Receptor ProbesNeerFamNo ratings yet
- Richard B. Rothman Et Al - Salvinorin A: Allosteric Interactions at The Mu-Opioid ReceptorDocument10 pagesRichard B. Rothman Et Al - Salvinorin A: Allosteric Interactions at The Mu-Opioid ReceptorNeerFamNo ratings yet
- Jeffrey L. Cummings and Bruce L. Miller - Visual Hallucinations: Clinical Occurrence and Use in Differential DiagnosisDocument6 pagesJeffrey L. Cummings and Bruce L. Miller - Visual Hallucinations: Clinical Occurrence and Use in Differential DiagnosisNeerFamNo ratings yet
- Michelle Gillis - Phenylethylamine: More Than Just A Pea-Sized NeurochemicalDocument3 pagesMichelle Gillis - Phenylethylamine: More Than Just A Pea-Sized NeurochemicalNeerFamNo ratings yet
- Amphetamines MechanismsDocument28 pagesAmphetamines Mechanisms50_BMGNo ratings yet
- Christina A. Schad Et Al - Endogenous Opioids in Dopaminergic Cell Body Regions Modulate Amphetamine-Induced Increases in Extracellular Dopamine Levels in The Terminal RegionsDocument7 pagesChristina A. Schad Et Al - Endogenous Opioids in Dopaminergic Cell Body Regions Modulate Amphetamine-Induced Increases in Extracellular Dopamine Levels in The Terminal RegionsNeerFamNo ratings yet
- Henry David Abraham Et Al - The Psychopharmacology of HallucinogensDocument14 pagesHenry David Abraham Et Al - The Psychopharmacology of HallucinogensNeerFamNo ratings yet
- Kyung-Ah Kim and Mark Von Zastrow - Old Drugs Learn New Tricks: Insights From Mammalian Trace Amine ReceptorsDocument3 pagesKyung-Ah Kim and Mark Von Zastrow - Old Drugs Learn New Tricks: Insights From Mammalian Trace Amine ReceptorsNeerFamNo ratings yet
- Richard T. Premont, Raul R. Gainetdinov and Marc G. Caron - Following The Trace of Elusive AminesDocument2 pagesRichard T. Premont, Raul R. Gainetdinov and Marc G. Caron - Following The Trace of Elusive AminesNeerFamNo ratings yet
- Kim K. Lemberg - The Pharmacology of Oxycodone: Studies in Vitro, in Vivo and in HumansDocument73 pagesKim K. Lemberg - The Pharmacology of Oxycodone: Studies in Vitro, in Vivo and in HumansNeerFamNo ratings yet
- Richard B. Herbert, Henrietta Venter and Sonja Pos - Do Mammals Make Their Own Morphine?Document6 pagesRichard B. Herbert, Henrietta Venter and Sonja Pos - Do Mammals Make Their Own Morphine?NeerFamNo ratings yet
- Endogenous Psychoactive Tryptamines Reconsidered An Anxiolytic Role For DimethyltryptamineDocument8 pagesEndogenous Psychoactive Tryptamines Reconsidered An Anxiolytic Role For DimethyltryptaminePedro S ReisNo ratings yet
- L.R. Watkins Et Al - Parallel Activation of Multiple Spinal Opiate Systems Appears To Mediate 'Non-Opiate' Stress-Induced AnalgesiasDocument10 pagesL.R. Watkins Et Al - Parallel Activation of Multiple Spinal Opiate Systems Appears To Mediate 'Non-Opiate' Stress-Induced AnalgesiasNeerFamNo ratings yet
- M. Johannessen Et Al - N, N-Dimethyltryptamine (DMT) As An Endogenous Ligand Candidate For The Sigma-1 ReceptorDocument1 pageM. Johannessen Et Al - N, N-Dimethyltryptamine (DMT) As An Endogenous Ligand Candidate For The Sigma-1 ReceptorNeerFamNo ratings yet
- George B. Stefano and Richard M. Kream - Endogenous Morphine Synthetic Pathway Preceded and Gave Rise To Catecholamine Synthesis in Evolution (Review)Document5 pagesGeorge B. Stefano and Richard M. Kream - Endogenous Morphine Synthetic Pathway Preceded and Gave Rise To Catecholamine Synthesis in Evolution (Review)PoloGreenNo ratings yet
- C.R.A. Leite-Panissi Et Al - Endogenous Opiate Analgesia Induced by Tonic Immobility in Guinea PigsDocument6 pagesC.R.A. Leite-Panissi Et Al - Endogenous Opiate Analgesia Induced by Tonic Immobility in Guinea PigsNeerFamNo ratings yet
- PNAS 2004 Poeaknapo 14091 6Document6 pagesPNAS 2004 Poeaknapo 14091 6Josué VelázquezNo ratings yet
- Wei Zhu, Kirk J. Mantione & George B. Stefano - Reticuline Exposure To Invertebrate Ganglia Increases Endogenous Morphine LevelsDocument8 pagesWei Zhu, Kirk J. Mantione & George B. Stefano - Reticuline Exposure To Invertebrate Ganglia Increases Endogenous Morphine LevelsNeerFamNo ratings yet
- Daniela Vigano, Tiziana Rubino and Daniela Parolaro - Molecular and Cellular Basis of Cannabinoid and Opioid InteractionsDocument9 pagesDaniela Vigano, Tiziana Rubino and Daniela Parolaro - Molecular and Cellular Basis of Cannabinoid and Opioid InteractionsNeerFamNo ratings yet
- Zuzana Justinova Et Al - The Endogenous Cannabinoid Anandamide and Its Synthetic Analog R (+) - Methanandamide Are Intravenously Self-Administered by Squirrel MonkeysDocument6 pagesZuzana Justinova Et Al - The Endogenous Cannabinoid Anandamide and Its Synthetic Analog R (+) - Methanandamide Are Intravenously Self-Administered by Squirrel MonkeysNeerFamNo ratings yet
- Emma Coddington Et Al - Endocannabinoids Mediate The Effects of Acute Stress and Corticosterone On Sex BehaviorDocument9 pagesEmma Coddington Et Al - Endocannabinoids Mediate The Effects of Acute Stress and Corticosterone On Sex BehaviorNeerFamNo ratings yet
- Fernando Rodriguez de Fonseca and Miriam Schneider - The Endogenous Cannabinoid System and Drug Addiction: 20 Years After The Discovery of The CB1 ReceptorDocument4 pagesFernando Rodriguez de Fonseca and Miriam Schneider - The Endogenous Cannabinoid System and Drug Addiction: 20 Years After The Discovery of The CB1 ReceptorNeerFamNo ratings yet
- David P. Finn - Endocannabinoid-Mediated Modulation of Stress Responses: Physiological and Pathophysiological SignificanceDocument58 pagesDavid P. Finn - Endocannabinoid-Mediated Modulation of Stress Responses: Physiological and Pathophysiological SignificanceNeerFamNo ratings yet
- Enza Palazzo - The Role of Cannabinoid Receptors in The Descending Modulation of PainDocument13 pagesEnza Palazzo - The Role of Cannabinoid Receptors in The Descending Modulation of PainNeerFamNo ratings yet
- A New Lingual Straight-Wire Techique: Journal of Clinical Orthodontics: JCO February 2010Document11 pagesA New Lingual Straight-Wire Techique: Journal of Clinical Orthodontics: JCO February 2010Hafaifa TaiebNo ratings yet
- Sodium Chloride (Rock Salt) MSDS: Section 1: Chemical Product and Company IdentificationDocument6 pagesSodium Chloride (Rock Salt) MSDS: Section 1: Chemical Product and Company IdentificationMohamed MaghawryNo ratings yet
- Anxiety Disorders BBDocument18 pagesAnxiety Disorders BBRyan Justin BoudreauxNo ratings yet
- Antidepression ReikiDocument6 pagesAntidepression ReikiDoc Lyman88% (8)
- Paket 2 OkDocument5 pagesPaket 2 OkBerna TodingNo ratings yet
- Intramedullary Nailing For Femur Fracture Web Version EnglishDocument6 pagesIntramedullary Nailing For Femur Fracture Web Version EnglishCarima JatnoNo ratings yet
- Basic Priciples of Heart SurgeryDocument6 pagesBasic Priciples of Heart SurgeryprofarmahNo ratings yet
- Vertigo MeniereDocument5 pagesVertigo MeniereHamba AllahNo ratings yet
- 1005 Hemorrhage ControlDocument88 pages1005 Hemorrhage ControlZainescu Dan100% (2)
- Jordan University of Science and TechnologyDocument33 pagesJordan University of Science and TechnologyNourAldin AbuSalehNo ratings yet
- DDFGGDocument2 pagesDDFGGstrafforNo ratings yet
- (2009) Study Guide To GeriatricPsychiatry - QsDocument41 pages(2009) Study Guide To GeriatricPsychiatry - QsPepe Garcia Estebez100% (1)
- Medical Surgical Nursing ReviewDocument97 pagesMedical Surgical Nursing ReviewBernadeth Josefa BorelaNo ratings yet
- 2015 Case 6 MBUDocument12 pages2015 Case 6 MBUDinesh TiwariNo ratings yet
- Impression Making in Complete DentureDocument128 pagesImpression Making in Complete DentureBharanijaNo ratings yet
- Medical Nutrition in Neurobehavioral2Document34 pagesMedical Nutrition in Neurobehavioral2Lia Dwi JayantiNo ratings yet
- Police Log November 7, 2016Document13 pagesPolice Log November 7, 2016MansfieldMAPoliceNo ratings yet
- Cassida Fluid GL 220-SdsDocument5 pagesCassida Fluid GL 220-SdsFajar EkoNo ratings yet
- Procalcitonin Testing To Guide Antibiotic Therapy in Acute Upper and Lower Respiratory Tract InfectionsDocument2 pagesProcalcitonin Testing To Guide Antibiotic Therapy in Acute Upper and Lower Respiratory Tract InfectionsMr. LNo ratings yet
- Fascio LaDocument20 pagesFascio LaMuhammad NoorNo ratings yet
- Trust Me, Im A (Junior) Doctor (Pemberton, Max (Pemberton, Max) )Document234 pagesTrust Me, Im A (Junior) Doctor (Pemberton, Max (Pemberton, Max) )Rakshith BalajiNo ratings yet
- CyberTherapy & Rehabilitation, Issue 3 (3), Winter 2010.Document52 pagesCyberTherapy & Rehabilitation, Issue 3 (3), Winter 2010.Giuseppe RivaNo ratings yet
- Interpretation of Histograms and Its Correlation WDocument5 pagesInterpretation of Histograms and Its Correlation WrezqiNo ratings yet
- Khalil High Yeild Step 2 Cs Mnemonic 2nd EdDocument24 pagesKhalil High Yeild Step 2 Cs Mnemonic 2nd EdCarolina Lopez100% (1)
- Anesthesia For TurpDocument5 pagesAnesthesia For Turptnim.dsNo ratings yet
- Immediate Care of The NewbornDocument4 pagesImmediate Care of The NewbornMichelle GambolNo ratings yet
- Diagnosis and Management of Subarachnoid HemorrhageDocument25 pagesDiagnosis and Management of Subarachnoid HemorrhageMisael ClintonNo ratings yet
- Dabur PresentationDocument26 pagesDabur PresentationShraddha Khurana100% (1)
- AA Interview Sou Fujimoto Andreas KoflerDocument2 pagesAA Interview Sou Fujimoto Andreas KoflerLong VuNo ratings yet