You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Emission Control SystemsDocument44 pagesEmission Control SystemsNilesh MeghwalNo ratings yet
- Suzuki DF2-5 Service ManualDocument148 pagesSuzuki DF2-5 Service ManualAnonymous PUNw29Xgr100% (1)
- Manual Motor Yamaha F115 PDFDocument98 pagesManual Motor Yamaha F115 PDFLitus Fendetestas100% (1)
- Mil-Std-3004d W-CH1 PDFDocument266 pagesMil-Std-3004d W-CH1 PDFPaul BarnardNo ratings yet
- Problem Set 1 - BM - SolutionsDocument16 pagesProblem Set 1 - BM - SolutionsSatrujit MohapatraNo ratings yet
- Cepmlp Car14 83 496182187Document19 pagesCepmlp Car14 83 496182187Antonio AulicinoNo ratings yet
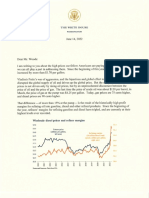
- Joe Biden Letter To Oil CompaniesDocument3 pagesJoe Biden Letter To Oil CompaniescharliespieringNo ratings yet
- Be General Owners ManualDocument68 pagesBe General Owners ManualMilburnDrysdaleNo ratings yet
- Operator's Manual: ERIES 1000Document40 pagesOperator's Manual: ERIES 1000Mark PhillipsNo ratings yet
- Jiangdong Motor Owner's ManualDocument14 pagesJiangdong Motor Owner's Manualrjcantwell67% (6)
- Motor Gasoline Specifications and Trend in IndiaDocument2 pagesMotor Gasoline Specifications and Trend in IndiavivekagNo ratings yet
- Gilera Stalker 50 - MaintenanceDocument56 pagesGilera Stalker 50 - MaintenanceMegan Read0% (1)
- Philippine Agricultural Engineers Board Review Pre-Exam QuestionsDocument12 pagesPhilippine Agricultural Engineers Board Review Pre-Exam QuestionsKen Lloyd F. Pabilan100% (4)
- 53 Icwai October Issue 2010Document80 pages53 Icwai October Issue 2010lotusvvNo ratings yet
- FuelDocument41 pagesFuelJasmine-Nezzar ImbigNo ratings yet
- Britelyt 500CP ManualDocument20 pagesBritelyt 500CP ManualtropascoutfranciscospallarossaNo ratings yet
- Business Project ShellDocument23 pagesBusiness Project ShellMuhammad Shafay MalikNo ratings yet
- Catalogue Mondea 2023 Fix (2) XDocument38 pagesCatalogue Mondea 2023 Fix (2) XyanurulNo ratings yet
- Bioenergy and BiofuelsDocument14 pagesBioenergy and BiofuelsA S M Younus Bhuiyan SabbirNo ratings yet
- Plunkett's Renewable, Alt. & Hydro. Energy Industry Almanac 2012 - Renewable, Alternative & Hydrogen Energy Industry Market Research, Statistics, Trends & Leading Companies (PDFDrive)Document514 pagesPlunkett's Renewable, Alt. & Hydro. Energy Industry Almanac 2012 - Renewable, Alternative & Hydrogen Energy Industry Market Research, Statistics, Trends & Leading Companies (PDFDrive)Ahmad ShamylNo ratings yet
- Tancet MBA: Quest I On Paper 2015Document23 pagesTancet MBA: Quest I On Paper 2015joyal JoyalNo ratings yet
- BP Tank Cleaning GuideDocument1 pageBP Tank Cleaning Guidesergey328No ratings yet
- Dual - Fuel - Genset - Engine CPCB PDFDocument59 pagesDual - Fuel - Genset - Engine CPCB PDFrakamechNo ratings yet
- Harmony II HRR 216-Sda PDFDocument72 pagesHarmony II HRR 216-Sda PDFacsscrufNo ratings yet
- General Information - Health and Safety PrecautionsDocument1,618 pagesGeneral Information - Health and Safety PrecautionsJose Augusto Melo100% (2)
- 3906 Qep Dec11Document88 pages3906 Qep Dec11ronfrendNo ratings yet
- DF25A - 30A Owner's ManualDocument77 pagesDF25A - 30A Owner's Manualshock2000No ratings yet
- Fuel Properties and TypesDocument67 pagesFuel Properties and TypesJagjot SinghNo ratings yet
- Supply Chain Management and E-Gas StationsDocument8 pagesSupply Chain Management and E-Gas StationsDivya AgarwalNo ratings yet