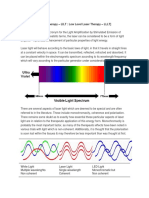
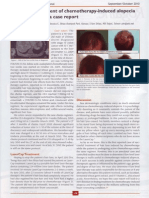
You might also like
- Stimulation and Transcranial Direct Current StimulationDocument453 pagesStimulation and Transcranial Direct Current StimulationMarcelo LugonNo ratings yet
- Microbiology With Diseases by Body System 5th Edition Bauman Test BankDocument25 pagesMicrobiology With Diseases by Body System 5th Edition Bauman Test BankRhondaHogancank100% (50)
- The Different Types of Electrotherapy DevicesDocument4 pagesThe Different Types of Electrotherapy DevicescassianohcNo ratings yet
- Transcranial Magnetic StimulationDocument2 pagesTranscranial Magnetic StimulationCandle Shop FullNo ratings yet
- Pulse Centers BrochureDocument22 pagesPulse Centers BrochureJohnNo ratings yet
- Processing of Bone SpecimenDocument100 pagesProcessing of Bone SpecimenAbhinav Arora0% (1)
- Epri Chemical Cleaning PDFDocument50 pagesEpri Chemical Cleaning PDFARSALAN GOPALNo ratings yet
- Temperature Control With ThermistorDocument7 pagesTemperature Control With ThermistorMonika Maheshwari100% (1)
- Tmecc 0411Document6 pagesTmecc 0411Karen BenteoNo ratings yet
- Effects of Contrast Sensitivity On Colour Vision TestingDocument33 pagesEffects of Contrast Sensitivity On Colour Vision TestingDorothy Eugene Nindya WiharyantoNo ratings yet
- The Cerebellum-8 PDFDocument83 pagesThe Cerebellum-8 PDFlovelots1234No ratings yet
- Does The Clock Drawing Test Have Focal Neuroanatomical CorrelatesDocument10 pagesDoes The Clock Drawing Test Have Focal Neuroanatomical CorrelatesIcaroNo ratings yet
- Fixatior Block GameDocument5 pagesFixatior Block GameperlatagNo ratings yet
- The History of Microcurrent Stimulation PDFDocument2 pagesThe History of Microcurrent Stimulation PDFjegm09100% (2)
- Brain Seizure Detection and Classification Using EEG SignalsFrom EverandBrain Seizure Detection and Classification Using EEG SignalsNo ratings yet
- Low Level Laser Therapy Clinical Report Philip OdegardDocument4 pagesLow Level Laser Therapy Clinical Report Philip OdegardLinda SwansonNo ratings yet
- The Use of Optokinetic Stimulation in VestibularDocument6 pagesThe Use of Optokinetic Stimulation in Vestibularbuku fisio0% (1)
- (Contemporary Clinical Neuroscience) Giuliana Grimaldi, Mario Manto (auth.), Giuliana Grimaldi, Mario Manto (eds.)-Mechanisms and Emerging Therapies in Tremor Disorders-Springer-Verlag New York (2013).pdfDocument490 pages(Contemporary Clinical Neuroscience) Giuliana Grimaldi, Mario Manto (auth.), Giuliana Grimaldi, Mario Manto (eds.)-Mechanisms and Emerging Therapies in Tremor Disorders-Springer-Verlag New York (2013).pdfDanalachi Adrian100% (2)
- Post Trauma Vision SyndromeDocument10 pagesPost Trauma Vision SyndromeEvelyn SepulvedaNo ratings yet
- LASER (LLLT) BASICS - "Mode of Action of Laser in Tissues"Document41 pagesLASER (LLLT) BASICS - "Mode of Action of Laser in Tissues"Dr. Mohammad Nazrul IslamNo ratings yet
- Carotid Artery Dissection PDFDocument3 pagesCarotid Artery Dissection PDFOdranoelRichamNo ratings yet
- PEMF and Exercises in Patients With Shoulder Impingement APMR 2014Document8 pagesPEMF and Exercises in Patients With Shoulder Impingement APMR 2014Michele GonçalvesNo ratings yet
- LightDocument84 pagesLightsenjicsNo ratings yet
- Mu Rythm in Children and SymptomsDocument94 pagesMu Rythm in Children and SymptomsAdrian GtNo ratings yet
- Neurostimulation For Stroke RehabilitationDocument11 pagesNeurostimulation For Stroke Rehabilitationisos.mporei.vevaios.No ratings yet
- Low-Level Laser Therapy by Philip OdegardDocument2 pagesLow-Level Laser Therapy by Philip OdegardLinda SwansonNo ratings yet
- Root Meristem, Root Cap, and RootDocument23 pagesRoot Meristem, Root Cap, and RootJulian ManongdoNo ratings yet
- Transcranial Magnetic Stimulation Power PointDocument32 pagesTranscranial Magnetic Stimulation Power PointDrGasnas50% (2)
- Brain Circuit Dysfunction in Post - Traumatic Stress Disorder: From Mouse To Man - Robert J. FensterDocument17 pagesBrain Circuit Dysfunction in Post - Traumatic Stress Disorder: From Mouse To Man - Robert J. FensterAlvaro Arboledas LoriteNo ratings yet
- Tesla S Inventions of Importance For MedDocument3 pagesTesla S Inventions of Importance For MedMorgan IonNo ratings yet
- QEEG Profile ReportDocument11 pagesQEEG Profile ReportAndrei BeraNo ratings yet
- Intraoperative Neurophysiological Monitoring 2011Document415 pagesIntraoperative Neurophysiological Monitoring 2011dr.darchNo ratings yet
- Eeg MigraineDocument20 pagesEeg MigraineAkhmad AfriantoNo ratings yet
- Light TherapyDocument32 pagesLight Therapylionrudrams81100% (1)
- Cerebral Blood Flow StrokeDocument122 pagesCerebral Blood Flow Strokeapi-3748748No ratings yet
- Fluorescent Light Full-SpectrumDocument15 pagesFluorescent Light Full-SpectrumAlexander WunschNo ratings yet
- Seminar On Dna Computing: Submitted To Miss - Bhawna Singhla N.C.C.EDocument20 pagesSeminar On Dna Computing: Submitted To Miss - Bhawna Singhla N.C.C.EAshish RanaNo ratings yet
- Six-Phase Table: International Society of HomotoxicologyDocument1 pageSix-Phase Table: International Society of HomotoxicologyPablo Matas SoriaNo ratings yet
- Blood Pressure and The Brain The Neurology of Hypertension PDFDocument12 pagesBlood Pressure and The Brain The Neurology of Hypertension PDFGustavo PestanaNo ratings yet
- Tinnitus PDFDocument10 pagesTinnitus PDFАлександра КондряNo ratings yet
- Transcranial Brain StimulationDocument456 pagesTranscranial Brain StimulationLakshmi Muralidhar67% (3)
- Cell Structure by Sabir.h.nDocument15 pagesCell Structure by Sabir.h.nprinceaimy.bestNo ratings yet
- Electroacupuncture According To Voll (EAV)Document1 pageElectroacupuncture According To Voll (EAV)vibe100No ratings yet
- Latest Hair Loss RESEARCH and TECHNIQUE Breakthrough in 2012Document8 pagesLatest Hair Loss RESEARCH and TECHNIQUE Breakthrough in 2012Dr Andrew KimNo ratings yet
- Noninvasive Human Brain Stimulation: Timothy Wagner, Antoni Valero-Cabre, and Alvaro Pascual-LeoneDocument41 pagesNoninvasive Human Brain Stimulation: Timothy Wagner, Antoni Valero-Cabre, and Alvaro Pascual-LeoneGhinea IulianaNo ratings yet
- Nerve Entrapment - UpdateDocument17 pagesNerve Entrapment - UpdatealobrienNo ratings yet
- Eyeball Motion Controlled Wheelchair Using IR SensorsDocument4 pagesEyeball Motion Controlled Wheelchair Using IR SensorssathishNo ratings yet
- Electricl StimulationDocument26 pagesElectricl StimulationSuvi AcesoNo ratings yet
- Richard Belli WorkshopDocument62 pagesRichard Belli WorkshopalicefreespiritNo ratings yet
- The Second Edition Expert Consensus Statements On The Use of IVIg in NeurologyDocument134 pagesThe Second Edition Expert Consensus Statements On The Use of IVIg in NeurologyRonald DewaNo ratings yet
- Hendri Saputra I11112043 Low-Level Laser Therapy in Medical RehabilitationDocument3 pagesHendri Saputra I11112043 Low-Level Laser Therapy in Medical RehabilitationHendri SaputraNo ratings yet
- Photobiological Fundamentals of Low-Power Laser TherapyDocument6 pagesPhotobiological Fundamentals of Low-Power Laser TherapyGabrielNo ratings yet
- Mechanisms and Applications of The Anti-Inflammatory Effects of PhotobiomodulationDocument25 pagesMechanisms and Applications of The Anti-Inflammatory Effects of PhotobiomodulationarexixNo ratings yet
- Lower Laser TherapyDocument17 pagesLower Laser TherapywahyuNo ratings yet
- Huang 2011Document17 pagesHuang 2011Brenda TakagiNo ratings yet
- Photosynthesis: Contents To Read in This ChapterDocument22 pagesPhotosynthesis: Contents To Read in This ChapterSajjad Ali ChadharNo ratings yet
- Photosynthesis: Contents To Read in This ChapterDocument22 pagesPhotosynthesis: Contents To Read in This ChapterSajjad Ali ChadharNo ratings yet
- Walt-low-Energy Laser Therapy As Quantum MedicineDocument4 pagesWalt-low-Energy Laser Therapy As Quantum Medicinemuitasorte9No ratings yet
- Laser TherapyDocument7 pagesLaser TherapyGustavo CabanasNo ratings yet
- Proposed Mechanisms of Photobiomodulation or Low-Level Light TherapyDocument37 pagesProposed Mechanisms of Photobiomodulation or Low-Level Light TherapySandra Morales OteroNo ratings yet
- Laser TherapyDocument7 pagesLaser TherapylecturioNo ratings yet
- Hamblin 2018Document14 pagesHamblin 2018Duccio Rossi Degl InnocentiNo ratings yet
- GrowToGive Participation Flyer - Congregation B'nai Israel Nov 4, 2016 Boca RatonDocument1 pageGrowToGive Participation Flyer - Congregation B'nai Israel Nov 4, 2016 Boca RatonAlan J Bauman MDNo ratings yet
- Simply The Best - Hair Transplant - DrAlanBauman MAR-APR 2017Document3 pagesSimply The Best - Hair Transplant - DrAlanBauman MAR-APR 2017Alan J Bauman MDNo ratings yet
- Invisible Baldness DrAlanBaumanDocument4 pagesInvisible Baldness DrAlanBaumanAlan J Bauman MDNo ratings yet
- Effect of Autologous Activated PRP Injection On Pattern Hair Loss - VCervelliDocument10 pagesEffect of Autologous Activated PRP Injection On Pattern Hair Loss - VCervelliAlan J Bauman MDNo ratings yet
- WCHR 2015 Final Conference Program NAHRSDocument48 pagesWCHR 2015 Final Conference Program NAHRSAlan J Bauman MD100% (1)
- Viviscal Marine Protein Safety-Efficacy - JDD-DrAlanBaumanDocument12 pagesViviscal Marine Protein Safety-Efficacy - JDD-DrAlanBaumanAlan J Bauman MDNo ratings yet
- PRP Platelet Rich Plasma Hair Growth - DrBauman - PrimeDocument7 pagesPRP Platelet Rich Plasma Hair Growth - DrBauman - PrimeAlan J Bauman MDNo ratings yet
- FDA Cleared LaserCap LCPRO K150613 Hair GrowthDocument7 pagesFDA Cleared LaserCap LCPRO K150613 Hair GrowthAlan J Bauman MDNo ratings yet
- PRP Platelet Rich Plasma FAQ - DR Alan BaumanDocument1 pagePRP Platelet Rich Plasma FAQ - DR Alan BaumanAlan J Bauman MDNo ratings yet
- Xelanj Tofacitinib Yale Alopecia Baldness Psoriasis Arthritis Cure Jid2014260a DrAlanBaumanDocument11 pagesXelanj Tofacitinib Yale Alopecia Baldness Psoriasis Arthritis Cure Jid2014260a DrAlanBaumanAlan J Bauman MDNo ratings yet
- ISHRS Program 2014 KualaLumpur Malaysia 2014 DrAlanBaumanDocument40 pagesISHRS Program 2014 KualaLumpur Malaysia 2014 DrAlanBaumanAlan J Bauman MDNo ratings yet
- ISHRS 21st Abstract Book 2013 SanFran DR Alan Bauman FinalDocument542 pagesISHRS 21st Abstract Book 2013 SanFran DR Alan Bauman FinalAlan J Bauman MDNo ratings yet
- Hair Loss Management PreConference Workshop A4M Orlando May 2014Document1 pageHair Loss Management PreConference Workshop A4M Orlando May 2014Alan J Bauman MDNo ratings yet
- Xeljanz-Tofacitinib FDA Label - DrAlanBaumanDocument27 pagesXeljanz-Tofacitinib FDA Label - DrAlanBaumanAlan J Bauman MDNo ratings yet
- Vampire PRP Hair Growth - South Florida Health Wellness - DR Alan BaumanDocument2 pagesVampire PRP Hair Growth - South Florida Health Wellness - DR Alan BaumanAlan J Bauman MD100% (1)
- Musical Cares - Sun Sentinal Article "Student of Note"Document1 pageMusical Cares - Sun Sentinal Article "Student of Note"Alan J Bauman MDNo ratings yet
- BaumanMD Therapeutic Anti-DHT Shampoo FAQDocument1 pageBaumanMD Therapeutic Anti-DHT Shampoo FAQAlan J Bauman MDNo ratings yet
- Xeljanz-Tofacitinib FDA Label - DrAlanBaumanDocument27 pagesXeljanz-Tofacitinib FDA Label - DrAlanBaumanAlan J Bauman MDNo ratings yet
- ISHRS 21st ProgramGuide 2013 SanFran DR Alan Bauman - FinalDocument52 pagesISHRS 21st ProgramGuide 2013 SanFran DR Alan Bauman - FinalAlan J Bauman MDNo ratings yet
- World Congress For Hair Research - WCHR 2013 AbstractsDocument49 pagesWorld Congress For Hair Research - WCHR 2013 AbstractsAlan J Bauman MDNo ratings yet
- 501 (C) 3 Approval For "Musical Cares"Document2 pages501 (C) 3 Approval For "Musical Cares"Alan J Bauman MDNo ratings yet
- LaserCap Use During Chemotherapy To Prevent Hair Loss - September/October 2010 - TK ShiaoDocument1 pageLaserCap Use During Chemotherapy To Prevent Hair Loss - September/October 2010 - TK ShiaoAlan J Bauman MDNo ratings yet
- 3b.boiler Treatment MethodsDocument76 pages3b.boiler Treatment Methodsalokbdas100% (1)
- RESONANCE Haloalkanes & HaloarenesDocument36 pagesRESONANCE Haloalkanes & HaloarenesAlok pandey67% (6)
- Introduction To Micro/Nano ElectronicsDocument51 pagesIntroduction To Micro/Nano ElectronicsLIAKMANNo ratings yet
- Cu ZN 10Document1 pageCu ZN 10Sara RiddleNo ratings yet
- Particulate Nature of MatterDocument4 pagesParticulate Nature of MatterSandy ItabNo ratings yet
- Duncan Sobre EsmaltesDocument9 pagesDuncan Sobre EsmaltesLeland StanfordNo ratings yet
- Polyaluminium Chloride: Product SpecificationsDocument2 pagesPolyaluminium Chloride: Product SpecificationsMonica Choi SeungjunhyungNo ratings yet
- Y. Osamura Et Al.: Rjournal of Molecular Structure Theochem 461 462 1999 399 416 400Document17 pagesY. Osamura Et Al.: Rjournal of Molecular Structure Theochem 461 462 1999 399 416 400IvanAgustínAymerichNo ratings yet
- KIMO HQ210 BrochureDocument4 pagesKIMO HQ210 BrochurebolsjhevikNo ratings yet
- Epoxy 2kDocument3 pagesEpoxy 2kMitra YuningsihNo ratings yet
- Numerical Analysis For Energy Performance Optimization of Hollow Bricks For Roofing. Case Study - Hot Climate of AlgeriaDocument10 pagesNumerical Analysis For Energy Performance Optimization of Hollow Bricks For Roofing. Case Study - Hot Climate of Algeriamah0809No ratings yet
- Elec6063 Optoelectronics and Lightwave Technology: Part 2: Leds and Lasers - 1 (Leds)Document43 pagesElec6063 Optoelectronics and Lightwave Technology: Part 2: Leds and Lasers - 1 (Leds)wanxin zhouNo ratings yet
- Reducing Eliminating ESD Hazards During PYRO OperationsDocument42 pagesReducing Eliminating ESD Hazards During PYRO OperationsAnonymous TI2bUTNo ratings yet
- Safety Data Sheet SDS For HIT-RE 500 V3 Epoxy Adhesive Documentation ASSET DOC LOC 5384987Document24 pagesSafety Data Sheet SDS For HIT-RE 500 V3 Epoxy Adhesive Documentation ASSET DOC LOC 5384987Librany RioNo ratings yet
- Comparative Evaluation of The Physicochemical and Sensorial Quality of Dragon Fruit Jams Made With Diferent Types of Citrus Juices As Acidulants - Turnitin SubmissionDocument59 pagesComparative Evaluation of The Physicochemical and Sensorial Quality of Dragon Fruit Jams Made With Diferent Types of Citrus Juices As Acidulants - Turnitin SubmissionCheng KaiNo ratings yet
- Fire Shield LFST - 42E: Premium Performance Industry Applications Product AvailabilityDocument1 pageFire Shield LFST - 42E: Premium Performance Industry Applications Product Availabilityray9699No ratings yet
- International Journal of Current Research in Biosciences and Plant BiologyDocument12 pagesInternational Journal of Current Research in Biosciences and Plant BiologyFrancisco C. Gonçalves GonçalvesNo ratings yet
- NTU MethodDocument15 pagesNTU MethodVenkatesh KandalamNo ratings yet
- Isolation of Caffeine From A Tea BagDocument7 pagesIsolation of Caffeine From A Tea BagShahriman Radzi67% (6)
- TBP, ASTM, and EFV ComparedDocument2 pagesTBP, ASTM, and EFV Comparedvicktorinox230388No ratings yet
- GENBIO2 - Lesson - The Central Dogma of Molecular BiologyDocument2 pagesGENBIO2 - Lesson - The Central Dogma of Molecular BiologyJazmaine SimbulanNo ratings yet
- Selection of Gas CompressorsDocument4 pagesSelection of Gas CompressorsDiegoNo ratings yet
- SorptionDocument44 pagesSorptionluckyprimeNo ratings yet
- GF-304CR Dossier.Document35 pagesGF-304CR Dossier.suria qaqcNo ratings yet
- Carbon Enrichment in Mo SteelDocument50 pagesCarbon Enrichment in Mo SteelDhananjay ShimpiNo ratings yet
- Chinook-Services-Wellsite GeologyDocument6 pagesChinook-Services-Wellsite Geologyincharge geologistNo ratings yet