You might also like
- Improving Ankle and Knee Joint Stability: Proprioceptive Balancefit Discs DrillsFrom EverandImproving Ankle and Knee Joint Stability: Proprioceptive Balancefit Discs DrillsNo ratings yet
- Degenerative Spine DiseaseDocument17 pagesDegenerative Spine DiseasekidrocksbodydblNo ratings yet
- Spinal StenosisDocument14 pagesSpinal Stenosisshwampa100% (3)
- Disk HerniationDocument27 pagesDisk HerniationDamiati78No ratings yet
- Neck & Back PainDocument232 pagesNeck & Back PainAatif_Saif_80No ratings yet
- Boys' Changing Voices - What We Know NowDocument14 pagesBoys' Changing Voices - What We Know NowNicholasNo ratings yet
- Spine Biomechanics: Mer/Bio Soft Tissue Mechanics SB-1Document11 pagesSpine Biomechanics: Mer/Bio Soft Tissue Mechanics SB-1Sarah Magdy100% (1)
- Morpho: Skeleton and Bone Reference Points: Anatomy for ArtistsFrom EverandMorpho: Skeleton and Bone Reference Points: Anatomy for ArtistsRating: 4.5 out of 5 stars4.5/5 (2)
- SOME Answers aren’t 100% Correct, So be ATTENTIONDocument48 pagesSOME Answers aren’t 100% Correct, So be ATTENTIONHussein Adam0% (1)
- The Skeletal SystemDocument5 pagesThe Skeletal Systemeducguide100% (1)
- Causes and Mechanism of Herniated Nucleus PulposusDocument14 pagesCauses and Mechanism of Herniated Nucleus PulposusJm Fernandez Gan RNNo ratings yet
- Applied Equine Nutriton and TrainingDocument233 pagesApplied Equine Nutriton and TrainingDragonMistress100% (1)
- Banton BiomechanicsDocument9 pagesBanton BiomechanicsIoan AndraNo ratings yet
- Herniated Nucleus Pulposus (Report) - 2Document27 pagesHerniated Nucleus Pulposus (Report) - 2Angelu Gabrielle CastroNo ratings yet
- TB SpineDocument15 pagesTB SpineChe AinNo ratings yet
- Intervertebral Disc)Document28 pagesIntervertebral Disc)arlinieNo ratings yet
- A Simple Guide to The Posture, Spine Diseases and Use in Disease DiagnosisFrom EverandA Simple Guide to The Posture, Spine Diseases and Use in Disease DiagnosisNo ratings yet
- Anatomy of the Lumbar SpineDocument39 pagesAnatomy of the Lumbar Spinepasha100% (1)
- Korean Hand Therapy - Dan Lobash LAcDocument13 pagesKorean Hand Therapy - Dan Lobash LActvmedicine100% (6)
- Prolapsed Disc SignsDocument14 pagesProlapsed Disc SignsMartin Sage100% (1)
- Spinal Curves ScoliosisDocument22 pagesSpinal Curves ScoliosisJennifer Mann100% (1)
- Cebu Doctors' University College of Nursing: Module 1F: Group WorksheetDocument55 pagesCebu Doctors' University College of Nursing: Module 1F: Group WorksheetPhilip Anthony FernandezNo ratings yet
- Continuum Spinal Cord AnatomyDocument25 pagesContinuum Spinal Cord AnatomyslojnotakNo ratings yet
- Goat Farming PDFDocument35 pagesGoat Farming PDFShiva KarthikNo ratings yet
- Classes of Vertebrates (Comparative Analysis)Document173 pagesClasses of Vertebrates (Comparative Analysis)Vinci EbordeNo ratings yet
- TB Spine - Laboratory Diagnosis, Management of TB Spine Including MDR CasesDocument61 pagesTB Spine - Laboratory Diagnosis, Management of TB Spine Including MDR CasesPramod N KNo ratings yet
- Cervical RadiculopathyDocument7 pagesCervical Radiculopathyrhymescsf100% (1)
- Sectional Anatomy - PET CT and SPECT CT (PDFDrive)Document473 pagesSectional Anatomy - PET CT and SPECT CT (PDFDrive)Mohammad AdnanNo ratings yet
- Anatomia de Los Discos Intervertebrales y FisiopatologiasDocument9 pagesAnatomia de Los Discos Intervertebrales y FisiopatologiasGabriela PerdumNo ratings yet
- Anatomi Patologi Diskus IntervetebralisDocument28 pagesAnatomi Patologi Diskus IntervetebralismhyamannanNo ratings yet
- Anatomy, Shoulder and Upper Limb, Clavicle - StatPearls - NCBI BookshelfDocument7 pagesAnatomy, Shoulder and Upper Limb, Clavicle - StatPearls - NCBI BookshelfRizqan Fahlevvi AkbarNo ratings yet
- Anatomical Aspects of Epidural and Spinal Analgesia: Review PaperDocument12 pagesAnatomical Aspects of Epidural and Spinal Analgesia: Review PaperAchmad MustikaNo ratings yet
- Disk DegenDocument25 pagesDisk DegenLaviniaBrănescuNo ratings yet
- AnatomyDocument9 pagesAnatomyannora daffaNo ratings yet
- Cervical Laminectomy: AnatomyDocument9 pagesCervical Laminectomy: AnatomyAgung Choro de ObesNo ratings yet
- Cervical Spondylosis PDFDocument46 pagesCervical Spondylosis PDFMadalinaTamasNo ratings yet
- Cervical SpondylosisDocument46 pagesCervical SpondylosisWalter Burton100% (1)
- Anatomy - Vertebral ColumnDocument5 pagesAnatomy - Vertebral ColumnArmelda Elda MuhollariNo ratings yet
- Thoracic vertebrae anatomy and functionsDocument4 pagesThoracic vertebrae anatomy and functionsNeirfla WassabiNo ratings yet
- Electrodiagnosis of Cervical RadiculopathyDocument12 pagesElectrodiagnosis of Cervical RadiculopathywutariNo ratings yet
- Nucleus Pulposus Herniation - Statpearls - Ncbi Bookshelf: June 2019Document8 pagesNucleus Pulposus Herniation - Statpearls - Ncbi Bookshelf: June 2019Ikhsan FebriansyahNo ratings yet
- Herniated Nucleus PulposusDocument12 pagesHerniated Nucleus PulposusZascha Fushiany GunawanNo ratings yet
- Intervertebral DiscDocument4 pagesIntervertebral DiscJcoNo ratings yet
- Lumbar Degenerative Disk Disease - StatPearls - NCBI BookshelfDocument6 pagesLumbar Degenerative Disk Disease - StatPearls - NCBI BookshelfAhmad SyahmiNo ratings yet
- Electrodiagnosisofcervical Radiculopathy: Kevin Hakimi,, David SpanierDocument12 pagesElectrodiagnosisofcervical Radiculopathy: Kevin Hakimi,, David Spanierkashif mansoorNo ratings yet
- Bogduk Meniscoide PDFDocument15 pagesBogduk Meniscoide PDFMatias Hassan LattzNo ratings yet
- Imaging Spinal Stenosis and Cervical MyelopathyDocument10 pagesImaging Spinal Stenosis and Cervical Myelopathysimona mariana dutuNo ratings yet
- IVDD Aging, Degeneration, and HerniationDocument10 pagesIVDD Aging, Degeneration, and HerniationFajrin Dwi SyaputraNo ratings yet
- Spinal Disc HerniationDocument5 pagesSpinal Disc HerniationJoshua Hong100% (1)
- Anatomical Variation of The Brachial PlexusDocument9 pagesAnatomical Variation of The Brachial Plexusjustinsanz77No ratings yet
- Lumbar Spine Stenosis: A Common Cause of Back and Leg Pain: Normal AnatomyDocument13 pagesLumbar Spine Stenosis: A Common Cause of Back and Leg Pain: Normal Anatomyhunter_axl01No ratings yet
- AD 5 Vertebral AnatomyDocument37 pagesAD 5 Vertebral AnatomyLintang ZerlinaNo ratings yet
- Nursing Care of The Patient Undergoing Lumbar Spinal Fusion: ReviewsDocument10 pagesNursing Care of The Patient Undergoing Lumbar Spinal Fusion: ReviewsMuammar100% (1)
- Thespinalcord: A Review of Functional NeuroanatomyDocument18 pagesThespinalcord: A Review of Functional NeuroanatomyasfwegereNo ratings yet
- Degenerative Disc Disease of The Spine - From Anatomy To Pathophysiology and Radiological Appearance With Morphological and Functional ConsiderationsDocument14 pagesDegenerative Disc Disease of The Spine - From Anatomy To Pathophysiology and Radiological Appearance With Morphological and Functional ConsiderationswenhsiaochuanNo ratings yet
- 01-Batra Et AlDocument14 pages01-Batra Et AlfajarvicNo ratings yet
- Okeson 2007Document19 pagesOkeson 2007Duy ChâuNo ratings yet
- Sacroiliac Joint Assessment - ExaminationDocument13 pagesSacroiliac Joint Assessment - ExaminationDr Reza ModarresNo ratings yet
- The Provocative Lumbar Facet JointDocument10 pagesThe Provocative Lumbar Facet JointfilipecorsairNo ratings yet
- Joints and Ligaments of VertebraeDocument7 pagesJoints and Ligaments of Vertebraerahma sajjadNo ratings yet
- Craniovertebral Junction Anatomy & RadiologyDocument130 pagesCraniovertebral Junction Anatomy & Radiologydrarunrao100% (1)
- Torticolis OdtDocument25 pagesTorticolis OdtAndreea CimpoiNo ratings yet
- Cervical Spondylosis PathophysiologyDocument5 pagesCervical Spondylosis PathophysiologyIkram HikmatyarNo ratings yet
- Definition of Shoulder DislocationDocument66 pagesDefinition of Shoulder DislocationDeepika verma100% (1)
- Lumbar Spine StenosisDocument12 pagesLumbar Spine StenosisParag DashatwarNo ratings yet
- Lec2 The Spine 2Document37 pagesLec2 The Spine 2sana mumtazNo ratings yet
- Herniated Nucleus PulposusDocument12 pagesHerniated Nucleus PulposusshezarNo ratings yet
- Herniated Nucleus PulposusDocument5 pagesHerniated Nucleus PulposusNestle FolienteNo ratings yet
- Vertebral Column and Spine AnatomyDocument86 pagesVertebral Column and Spine AnatomyHà Quang SơnNo ratings yet
- Cervical Spondylosis: CordDocument4 pagesCervical Spondylosis: CordMiftahul Jannah GafarNo ratings yet
- Spinal Stenosis and Neurogenic Claudication - StatPearls - NCBI BookshelfDocument8 pagesSpinal Stenosis and Neurogenic Claudication - StatPearls - NCBI BookshelfchhabraanNo ratings yet
- Braddoms 2020 - Capitulo 13 - Ortesis EspinalesDocument13 pagesBraddoms 2020 - Capitulo 13 - Ortesis EspinalesSergio Navarrete VidalNo ratings yet
- Treatment of Cervical Spondylotic MyelopathyDocument49 pagesTreatment of Cervical Spondylotic MyelopathyencrownterNo ratings yet
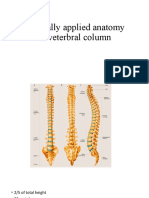
- Clinically Applied Anatomy of Veterbral ColumnDocument25 pagesClinically Applied Anatomy of Veterbral ColumnNiroshan VadivelNo ratings yet
- C1 Atlas C2 Axis C3 - C6 Typical Cervical Vertebra C7 Vertebra ProminensDocument4 pagesC1 Atlas C2 Axis C3 - C6 Typical Cervical Vertebra C7 Vertebra ProminensmiallyannaNo ratings yet
- 6.S, R, U Systems of Female PelvisDocument2 pages6.S, R, U Systems of Female Pelvisahmned A DOCTORNo ratings yet
- Cloward's ProcedureDocument30 pagesCloward's ProcedureDr.Humayun Rashid SagorNo ratings yet
- UsrDocument80 pagesUsrAntony QuispeNo ratings yet
- Human Anatomy Assignment 1. DR Onanuga-2021Document7 pagesHuman Anatomy Assignment 1. DR Onanuga-2021AlexanderNo ratings yet
- Flexicurve - Protocol: The Measure ExplainedDocument2 pagesFlexicurve - Protocol: The Measure ExplainedRaihan AdamNo ratings yet
- Human Skeleton Concept MapDocument1 pageHuman Skeleton Concept MapSalil ShauNo ratings yet
- Biomechanics of SpineDocument9 pagesBiomechanics of SpineAdosotoNo ratings yet
- Back Muscles - PhysiopediaDocument1 pageBack Muscles - PhysiopediaAline SánchezNo ratings yet
- OsteologyDocument96 pagesOsteologyBenita Putri MD100% (2)
- Nsci280 Nsci 280Document75 pagesNsci280 Nsci 280NSCI 280No ratings yet
- Nyala Scales DAKDocument6 pagesNyala Scales DAKLaura RicoNo ratings yet
- Treating Discus Hernia Using A Self Developed Method - A New Technology Without Surgical InterventionDocument6 pagesTreating Discus Hernia Using A Self Developed Method - A New Technology Without Surgical InterventionlazarstosicNo ratings yet
- Pediatric Spinal Schwannoma Case ReportDocument17 pagesPediatric Spinal Schwannoma Case ReportrakeNo ratings yet
- Anatomy of Vertebral ColumnDocument59 pagesAnatomy of Vertebral ColumnRivan DwiutomoNo ratings yet