You might also like
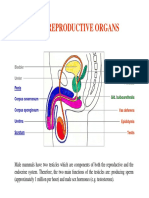
- 11 MaleDocument14 pages11 MaleJasmine KaurNo ratings yet
- Human Chromosomes: An Illustrated Introduction to Human CytogeneticsFrom EverandHuman Chromosomes: An Illustrated Introduction to Human CytogeneticsRating: 5 out of 5 stars5/5 (1)
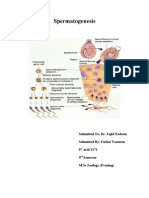
- Sajad 2020Document22 pagesSajad 2020MD LarasatiNo ratings yet
- Biology Investigatory Project On GametogenesisDocument20 pagesBiology Investigatory Project On GametogenesisAshwin Rockzz46% (13)
- Spermatogenesis - Biology Science Fair Project IdeasDocument8 pagesSpermatogenesis - Biology Science Fair Project IdeasEPFO AKURDI100% (1)
- Department of Morphology: Penza State University Medical InstituteDocument18 pagesDepartment of Morphology: Penza State University Medical InstituteunnatiNo ratings yet
- Spermatogenesis WPS Office 1Document12 pagesSpermatogenesis WPS Office 1nb6pd5rvnxNo ratings yet
- Histology of The Male Reproductive SystemDocument18 pagesHistology of The Male Reproductive SystemMuhammad JameelNo ratings yet
- Kfo181 Maennl GoDocument18 pagesKfo181 Maennl Gosong lyrics-sing your ownNo ratings yet
- Regeneration in AxolotlDocument13 pagesRegeneration in AxolotlȺនђᎥនђ ツᦔøץNo ratings yet
- Spermatogenesis:: (1) Formation of TheDocument3 pagesSpermatogenesis:: (1) Formation of ThebarbacumlaudeNo ratings yet
- Male Reproductive PhysiologyDocument83 pagesMale Reproductive PhysiologyX Christian CatalanNo ratings yet
- Sertoli Cell Cell Testes Germ Cells SertoliDocument3 pagesSertoli Cell Cell Testes Germ Cells Sertolihur-azmi-1154No ratings yet
- SPERMATOGENESISDocument58 pagesSPERMATOGENESISkathyNo ratings yet
- CC-13 Unit 1Document10 pagesCC-13 Unit 1Subhradeep SinghaNo ratings yet
- Male R.S. Dr Hekmat هاند اوتDocument15 pagesMale R.S. Dr Hekmat هاند اوتJo adamNo ratings yet
- Malereproductive (Author T.globa)Document40 pagesMalereproductive (Author T.globa)Cristina caimac100% (1)
- New Jyothi Central School Senior Secondary: CertificateDocument17 pagesNew Jyothi Central School Senior Secondary: CertificateSameeth Raj.K.SNo ratings yet
- Spermatogenesis: Feriha Ercan, Ph.D. Dept. Histology and EmbryologyDocument41 pagesSpermatogenesis: Feriha Ercan, Ph.D. Dept. Histology and EmbryologyErdem AltunNo ratings yet
- SpermatogenesisDocument13 pagesSpermatogenesiszuha khanNo ratings yet
- Spermatogenesis: Spermatogenesis Is The Process by Which MaleDocument6 pagesSpermatogenesis: Spermatogenesis Is The Process by Which MaleHusni AndiNo ratings yet
- Spermatogenesis: Created by Aji SantikoDocument16 pagesSpermatogenesis: Created by Aji SantikoPrince of Darkness100% (1)
- Reproduction Student MaterialDocument27 pagesReproduction Student MaterialEsosa OdighizuwaNo ratings yet
- Male Reproductive SytemDocument4 pagesMale Reproductive SytemSheena Liegh EntredichoNo ratings yet
- Lecture Notes Module - 3 - Male Reproductive Anatomy and PhysiologyDocument8 pagesLecture Notes Module - 3 - Male Reproductive Anatomy and PhysiologyMariaImranNo ratings yet
- Hormonal Control of GametogenesisDocument29 pagesHormonal Control of GametogenesisWachiel Arhamz100% (2)
- GametogenesisDocument35 pagesGametogenesisJason TulipatNo ratings yet
- SPERMATOGENESIS Spermatids Zirkin and Goldberg 2018Document5 pagesSPERMATOGENESIS Spermatids Zirkin and Goldberg 2018LudimilaNo ratings yet
- HD 201 E1 20140127 Histology of The Male Reproductive SystemDocument7 pagesHD 201 E1 20140127 Histology of The Male Reproductive SystemMaxine Alba100% (1)
- SpermatogenesisDocument8 pagesSpermatogenesisapi-26413035No ratings yet
- LAB EXERCISE 1 - SpermatogenesisDocument5 pagesLAB EXERCISE 1 - SpermatogenesisGerald Angelo DeguinioNo ratings yet
- CLASS 12 BIO PROJECT SpermatogenesisDocument19 pagesCLASS 12 BIO PROJECT Spermatogenesisanon_38352456389% (9)
- CLASS 12 BIO PROJECT SpermatogenesisDocument19 pagesCLASS 12 BIO PROJECT Spermatogenesisanon_383524563No ratings yet
- 24 Male Reproduction NXPowerLiteDocument141 pages24 Male Reproduction NXPowerLiteEarn cruzNo ratings yet
- 61 Sharma and Agarwal Spermiogenesis - An OverviewDocument26 pages61 Sharma and Agarwal Spermiogenesis - An Overviewjohairah merphaNo ratings yet
- Meiosis Is The First Step in Gametogenesis: Separation of Homologous Chromosomes Into Haploid Daughter CellsDocument18 pagesMeiosis Is The First Step in Gametogenesis: Separation of Homologous Chromosomes Into Haploid Daughter CellsMichael DiosesNo ratings yet
- Kuliah 2. Dasar Biomolekuler Reproduksi PriaDocument47 pagesKuliah 2. Dasar Biomolekuler Reproduksi PriaSeptiaNo ratings yet
- TestesDocument39 pagesTestesImadShaheen50% (2)
- The Male Genital SystemDocument5 pagesThe Male Genital SystemDr-AymanBahgatNo ratings yet
- YashDocument7 pagesYashShrutik DhongadeNo ratings yet
- EMBRYOLOGY: Gametogenesis, Fertilization in Details.Document46 pagesEMBRYOLOGY: Gametogenesis, Fertilization in Details.Mercy AdeolaNo ratings yet
- 26.4C: Spermatogenesis: Learning Objective Key Takeaways Key PointsDocument3 pages26.4C: Spermatogenesis: Learning Objective Key Takeaways Key Pointsmuhammad zeeshan ansarNo ratings yet
- Spermatogenesis: An Overview: Rakesh Sharma and Ashok AgarwalDocument25 pagesSpermatogenesis: An Overview: Rakesh Sharma and Ashok Agarwalumar umarNo ratings yet
- The Reproductive SystemDocument68 pagesThe Reproductive SystemSamuelNo ratings yet
- Fertilizacion RevisionDocument8 pagesFertilizacion RevisionAsistente IRP RESTREPONo ratings yet
- Embryology 3Document64 pagesEmbryology 3wasifuddin464No ratings yet
- Reproductive SystemDocument61 pagesReproductive SystemHetvi PatelNo ratings yet
- Gametogenesis: Gametogenesis Is The Process by Which Diploid Precursor Cells Undergo MeioticDocument2 pagesGametogenesis: Gametogenesis Is The Process by Which Diploid Precursor Cells Undergo MeioticnolaNo ratings yet
- SpermatogenesisDocument27 pagesSpermatogenesisSabio DenmenNo ratings yet
- Devl Biol-SpermatogenesisDocument9 pagesDevl Biol-SpermatogenesisSuresh DavuluriNo ratings yet
- Gametogenesis: by Anagha Jose Class - Xii Reg ScienceDocument16 pagesGametogenesis: by Anagha Jose Class - Xii Reg ScienceanaghaNo ratings yet
- Gametogenesis by Anagha JoseDocument16 pagesGametogenesis by Anagha JoseanaghaNo ratings yet
- Reproduction 1Document79 pagesReproduction 1AyeshaNo ratings yet
- Male Reproductive SystemDocument62 pagesMale Reproductive SystemSalman KhanNo ratings yet
- Gametogenesis Process in HumanDocument10 pagesGametogenesis Process in HumanFerhaeeza KalayakanNo ratings yet
- Lesson Two - Reproduction in MammalsDocument5 pagesLesson Two - Reproduction in MammalsNobody JamesNo ratings yet
- Gametogenesis, Gamete Transport & FertilizationDocument56 pagesGametogenesis, Gamete Transport & FertilizationvictorNo ratings yet
- Testis Physiology-Overview and Histology: January 2018Document13 pagesTestis Physiology-Overview and Histology: January 2018Sephendra LutfiNo ratings yet
- Spermatogenesis and OogenesisDocument17 pagesSpermatogenesis and Oogenesisanju kumawatNo ratings yet
- Grandiloquent DictionaryDocument116 pagesGrandiloquent DictionarykelamistgirisimNo ratings yet
- 5TH SemDocument1 page5TH SemKOTAMRAJUNo ratings yet
- Cyber SpaceDocument10 pagesCyber SpaceKOTAMRAJUNo ratings yet
- Constitution of India-Complete Full TextDocument471 pagesConstitution of India-Complete Full TextDaras Bir Singh67% (6)
- How To Build Muscle On A Raw Food DietDocument35 pagesHow To Build Muscle On A Raw Food DietTiago Matos100% (2)
- The Earliest Relationship Parents, Infants, and The Drama of EaDocument280 pagesThe Earliest Relationship Parents, Infants, and The Drama of EaBogdan BNo ratings yet
- Acne and Rosacea - Goldberg, David J, Berlin, AlexanderDocument98 pagesAcne and Rosacea - Goldberg, David J, Berlin, AlexanderAjayAsthanaNo ratings yet
- Androgens in Women: Androgen-Mediated Skin Disease and Patient EvaluationDocument10 pagesAndrogens in Women: Androgen-Mediated Skin Disease and Patient EvaluationPaulo Victor Amorim MarquesNo ratings yet
- Chemical Wizardry CompleteDocument132 pagesChemical Wizardry CompleteSaeed Khushal100% (5)
- From Pathogenesis of Acne Vulgaris To Anti-Acne AgentsDocument13 pagesFrom Pathogenesis of Acne Vulgaris To Anti-Acne AgentsNazihan Safitri AlkatiriNo ratings yet
- 2.the Nursing Role in Reproductive and Sexual HealthDocument70 pages2.the Nursing Role in Reproductive and Sexual HealthAngel Gabriel FornillosNo ratings yet
- Nursing Pharmacology Study GuideDocument53 pagesNursing Pharmacology Study Guidezsazsageorge100% (5)
- Anabolic SteriodsDocument43 pagesAnabolic SteriodsAhmed Safi100% (1)
- Instructional Material For Gender - FINALDocument64 pagesInstructional Material For Gender - FINALBenzDave AgsoyNo ratings yet
- SteroidsFromPhysiology2ClinicalMedicineITO12 PDFDocument220 pagesSteroidsFromPhysiology2ClinicalMedicineITO12 PDFJasonNo ratings yet
- Phytoestrogen TilapiaDocument7 pagesPhytoestrogen TilapiaAbdel-Fattah M. El-SayedNo ratings yet
- Pathology and Medical Therapy of Benign Prostatic HyperplasiaDocument5 pagesPathology and Medical Therapy of Benign Prostatic HyperplasiaRose Deasy100% (1)
- Shinjita Das M.D., Rachel Reynolds M.D. (Auth.), Joshua Zeichner (Eds.) - Acneiform Eruptions in Dermatology - A Differential Diagnosis-Springer-Verlag New York (2014) PDFDocument405 pagesShinjita Das M.D., Rachel Reynolds M.D. (Auth.), Joshua Zeichner (Eds.) - Acneiform Eruptions in Dermatology - A Differential Diagnosis-Springer-Verlag New York (2014) PDFMaryNo ratings yet
- Reproductive PhysiologyDocument40 pagesReproductive PhysiologyBaiq Trisna Satriana100% (1)
- Bodybuilding Nutrition 4 - Growth Hormone BoostersDocument10 pagesBodybuilding Nutrition 4 - Growth Hormone BoostersKINETO62100% (1)
- Biological Basis of HomosexulaityDocument22 pagesBiological Basis of HomosexulaityDhimitri BibolliNo ratings yet
- Boys' Behavior: Why Boys Behave The Way They DoDocument7 pagesBoys' Behavior: Why Boys Behave The Way They DokokoriNo ratings yet
- Placenta 2Document21 pagesPlacenta 2Nurba Enda Karina NasutionNo ratings yet
- DR Kyle Gillett Tools For Hormone Optimization in MalesDocument56 pagesDR Kyle Gillett Tools For Hormone Optimization in Maleswaheed khanNo ratings yet
- Steroid Half Life'sDocument6 pagesSteroid Half Life'sTomPark1No ratings yet
- Andropause A Review of The Definition and TreatmentDocument6 pagesAndropause A Review of The Definition and TreatmentJesus EtNo ratings yet
- Disorders of The Ovaries 2Document92 pagesDisorders of The Ovaries 2rerenNo ratings yet
- IJPC 14 2 Compounding For ManDocument92 pagesIJPC 14 2 Compounding For ManMatiasNo ratings yet
- HypospadiaDocument11 pagesHypospadiaweniNo ratings yet
- Hormonal Influences On Human Behavior PDFDocument22 pagesHormonal Influences On Human Behavior PDFKILAMAZUKE0% (1)
- Development of The Gental SystemDocument24 pagesDevelopment of The Gental SystemAustine OdhiamboNo ratings yet
- Puberty and The HPG AxisDocument36 pagesPuberty and The HPG AxiskjhkaNo ratings yet
- Dark, Firm and Dry Beef-Lectura 3Document4 pagesDark, Firm and Dry Beef-Lectura 3Andrea Pilar Ramos RossiNo ratings yet
- Christian Thibaudeau DR Jekyll MR Hyde PDFDocument200 pagesChristian Thibaudeau DR Jekyll MR Hyde PDFPauloCarvalho100% (3)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (378)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (516)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (6)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (15)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemFrom EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemRating: 4.5 out of 5 stars4.5/5 (115)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- Seven and a Half Lessons About the BrainFrom EverandSeven and a Half Lessons About the BrainRating: 4 out of 5 stars4/5 (109)
- Lymph & Longevity: The Untapped Secret to HealthFrom EverandLymph & Longevity: The Untapped Secret to HealthRating: 4.5 out of 5 stars4.5/5 (13)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldFrom EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldRating: 4 out of 5 stars4/5 (595)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- Change Your Brain, Change Your Life (Before 25): Change Your Developing Mind for Real-World SuccessFrom EverandChange Your Brain, Change Your Life (Before 25): Change Your Developing Mind for Real-World SuccessRating: 4 out of 5 stars4/5 (18)