You might also like
- Anatomie Humaine: Un Manuel Pratique et Intuitif pour Découvrir le Corps Humain et Toutes ses ComposantesFrom EverandAnatomie Humaine: Un Manuel Pratique et Intuitif pour Découvrir le Corps Humain et Toutes ses ComposantesNo ratings yet
- Développement EmbryonnaireDocument9 pagesDéveloppement EmbryonnaireEva Peignier100% (1)
- Cours Biologie Animale SNV l1 2015 Introduction GeneraleDocument4 pagesCours Biologie Animale SNV l1 2015 Introduction GeneraleSabrin Amani100% (1)
- CafDocument86 pagesCafLàbÿôuï ÃbderráhmàñëNo ratings yet
- Biologie Animale 3eme Trimestre PDFDocument29 pagesBiologie Animale 3eme Trimestre PDFFafi AliNo ratings yet
- Biologie Du Dévellopement (Synthèse)Document30 pagesBiologie Du Dévellopement (Synthèse)Nicoleta ChivuNo ratings yet
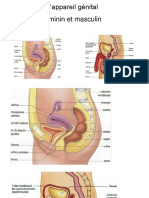
- 3 APPAREIL GENITAL FEMININ WordDocument13 pages3 APPAREIL GENITAL FEMININ WordFaïsole Adimou Ola100% (2)
- Transmission Des CaractèresDocument36 pagesTransmission Des CaractèresYaya TraoréNo ratings yet
- Embryologie HumaineDocument45 pagesEmbryologie Humainejeanmarctang76No ratings yet
- Anatomie de L Appareil Genital FinalDocument10 pagesAnatomie de L Appareil Genital Finalشكوب ستان100% (1)
- 03 OvogeneseDocument62 pages03 Ovogenesedjamila benaissaNo ratings yet
- Gamatogenese Et FecondationDocument47 pagesGamatogenese Et FecondationAmïn Fïlalï100% (1)
- TD N5. Exempl Corr.Document8 pagesTD N5. Exempl Corr.Mehdi Si-HassenNo ratings yet
- La MeioseDocument4 pagesLa MeioseKhaoulaFaithfulNo ratings yet
- Cours Rénale L2Document72 pagesCours Rénale L2ŚpĕĈíał ØñeNo ratings yet
- Physiologie Générale 2005, Le SyllabusDocument167 pagesPhysiologie Générale 2005, Le SyllabusAnou Kalenga100% (1)
- QE D Embryologie RéponseDocument11 pagesQE D Embryologie RéponseasmaeNo ratings yet
- 11-Introduction À L'helminthologieDocument47 pages11-Introduction À L'helminthologieYahia BenaliNo ratings yet
- Embryologie Humaine Polycopie PDF 1Document111 pagesEmbryologie Humaine Polycopie PDF 1LAFAYNo ratings yet
- CF NématodesDocument14 pagesCF NématodesalmnaouarNo ratings yet
- Embryologie Du Systeme Uro-GenitalDocument44 pagesEmbryologie Du Systeme Uro-Genitalsaidbenmokhtar3960No ratings yet
- Appareil Génital MaleDocument49 pagesAppareil Génital MaleTreycia PamboNo ratings yet
- 6eme Séance - 2Document42 pages6eme Séance - 2Mohamed AlouaneNo ratings yet
- TP Hématologie GénéraleDocument38 pagesTP Hématologie Généralemanarkhatib905No ratings yet
- Croissance CellulaireDocument28 pagesCroissance Cellulaireهوبة سفيلNo ratings yet
- Cours D'embryologie Spéciale G3 BMDocument220 pagesCours D'embryologie Spéciale G3 BMChristine Kosi100% (1)
- Tle D SVTEEHBDocument6 pagesTle D SVTEEHBJOEL LOIC100% (1)
- HématopoéseDocument38 pagesHématopoéseAmina BouzidiNo ratings yet
- Anatomie Physiologie SF S1Document7 pagesAnatomie Physiologie SF S1Rayhane Ben SaadaNo ratings yet
- L'appareil ReproducteurDocument118 pagesL'appareil ReproducteurAlexiaFoderaNo ratings yet
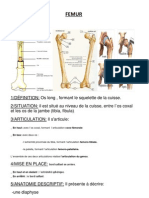
- FemurDocument4 pagesFemurBesma SbNo ratings yet
- Chapitre III La FécondationDocument7 pagesChapitre III La FécondationDel HadNo ratings yet
- 03 LE GLOBULE ROUGE Cours 4eme Année Pharmacie DR ABBASSENDocument4 pages03 LE GLOBULE ROUGE Cours 4eme Année Pharmacie DR ABBASSENAziz BenachourNo ratings yet
- Parasito de Blida PDFDocument104 pagesParasito de Blida PDFstimofteiNo ratings yet
- 12-Les Cestodes AdultesDocument68 pages12-Les Cestodes AdultesYahia BenaliNo ratings yet
- Apparei L Genital Misa JourDocument21 pagesApparei L Genital Misa JourBakary SanogoNo ratings yet
- Trichocephalose L3Document35 pagesTrichocephalose L3Sahouin Claude MedardNo ratings yet
- Méthode D'étude de La CelluleDocument33 pagesMéthode D'étude de La CelluleliliNo ratings yet
- Cours Axe Hypothalamo HypophysaireDocument51 pagesCours Axe Hypothalamo HypophysaireSamar OmraniNo ratings yet
- FécondationDocument47 pagesFécondationEric Charles HadjiNo ratings yet
- 5 Les Annexes EmbryonnairesDocument12 pages5 Les Annexes EmbryonnairesBook Hunter100% (2)
- Correction TD Bacteriologie Seance 1Document32 pagesCorrection TD Bacteriologie Seance 1Alexandre Kpangny BéniNo ratings yet
- Unité 1 - La Cellule, Unité Du VivantDocument8 pagesUnité 1 - La Cellule, Unité Du VivantIrene PGNo ratings yet
- LA GROSSESSE CorrigéDocument5 pagesLA GROSSESSE CorrigéGabriel UguenNo ratings yet
- Cours de Biochimie IdeDocument14 pagesCours de Biochimie IdeDessenbe TheophileNo ratings yet
- Ifsi Anatomie Physiologie Appareil LocomoteurDocument80 pagesIfsi Anatomie Physiologie Appareil LocomoteurZoulkiffiNo ratings yet
- LIPIDES Biochimie StructuraleDocument82 pagesLIPIDES Biochimie StructuraleCheick SANOUNo ratings yet
- Cours Endocrinologie de La ReproductionDocument49 pagesCours Endocrinologie de La ReproductionLukusa MauriceNo ratings yet
- TD Respiration 1Document6 pagesTD Respiration 1Busha100% (1)
- 6 - ProstaglandinesDocument10 pages6 - Prostaglandinesdjamila benaissaNo ratings yet
- 9-Généralités Sur Les HelminthesDocument9 pages9-Généralités Sur Les HelminthesBMA belguedjNo ratings yet
- Embryologie DescriptiveDocument5 pagesEmbryologie Descriptiveskeleton93No ratings yet
- Sol D Divisions CellDocument6 pagesSol D Divisions CellNana Diakité100% (1)
- Plathelminthes Némathelminthes PowerPointDocument44 pagesPlathelminthes Némathelminthes PowerPointBejen BEJENNo ratings yet
- INFERTILITÉ I. DÉFINITION II. LES FACTEURS QUI INFLUENCENT LA FERTILITÉ. 1 Age. 2 Maladies GénétiquesDocument24 pagesINFERTILITÉ I. DÉFINITION II. LES FACTEURS QUI INFLUENCENT LA FERTILITÉ. 1 Age. 2 Maladies GénétiquesOum BanatNo ratings yet
- Reproduction Humaine Fiche RevisionDocument2 pagesReproduction Humaine Fiche Revisionrayene chelly100% (1)
- Production de Proteines RecombinantesDocument96 pagesProduction de Proteines RecombinantesSaid Bellahcen100% (1)
- Tableau Des Types OeufDocument1 pageTableau Des Types Oeufraouf teach 04No ratings yet
- TD GénétiqueDocument4 pagesTD GénétiqueRaka Blue100% (2)
- Coqueluche 11 12Document2 pagesCoqueluche 11 12andreea.herescuNo ratings yet
- Une Nouvelle Classification en Parodontologie (Chicago 2017) - Impact en OrthodontieDocument6 pagesUne Nouvelle Classification en Parodontologie (Chicago 2017) - Impact en OrthodontieLilia RebaiNo ratings yet
- PLURIGIN Ovuli Foglietto Illustrativo Rev.30.10.18 (Francese)Document1 pagePLURIGIN Ovuli Foglietto Illustrativo Rev.30.10.18 (Francese)Magdalena LauraNo ratings yet
- ch14 241 LDPDocument6 pagesch14 241 LDPImaneNo ratings yet
- 5.1.4. Qualité Microbiologique Des Préparations Pharmaceutiques Et Des Substances Pour Usage Pharmaceutique Non StérilesDocument2 pages5.1.4. Qualité Microbiologique Des Préparations Pharmaceutiques Et Des Substances Pour Usage Pharmaceutique Non StérilesABDO ELJA100% (1)
- 10-Bases Immunologiques de La VaccinationDocument39 pages10-Bases Immunologiques de La VaccinationBMA belguedjNo ratings yet
- Directives-2021 PEC Palu VF 26-.4-2022Document51 pagesDirectives-2021 PEC Palu VF 26-.4-2022Joseph Ouoba100% (1)
- Manuel Indicateurs Secteur Sant - Final CSBDocument11 pagesManuel Indicateurs Secteur Sant - Final CSBANDRIAMIHAJASON Diamondra100% (1)
- Insuffisance Rénale Aigue: Préparer Par Guemmoula MarouaDocument29 pagesInsuffisance Rénale Aigue: Préparer Par Guemmoula Marouajames100% (1)
- Chap4 Pepiniere Et Plantation-1Document13 pagesChap4 Pepiniere Et Plantation-1Venite GeorgesNo ratings yet
- Normes Et Directives Chlorhexidine Ocytocyne Misoprostol Cyrille Du 13 06 2015 VFDocument23 pagesNormes Et Directives Chlorhexidine Ocytocyne Misoprostol Cyrille Du 13 06 2015 VFkadjungaNo ratings yet
- Manifestations Ophtalmiques Orbitopalpebrales de L'obstruction Nasale. Di R. Gola Et Al., 13 09 15Document14 pagesManifestations Ophtalmiques Orbitopalpebrales de L'obstruction Nasale. Di R. Gola Et Al., 13 09 15Anders CroccoNo ratings yet
- FQSDFDocument5 pagesFQSDFalexNo ratings yet
- La Checklist de La Médecine VasculaireDocument304 pagesLa Checklist de La Médecine Vasculairemaroua aouadi100% (1)
- 2021 04 20 15 23 37 RésultatsDocument2 pages2021 04 20 15 23 37 RésultatsImraan MoussaNo ratings yet
- GrossesseDocument18 pagesGrossessefullmetalporo2012No ratings yet
- Alchimie Paracelse (1493-1541) - de La Teinture Des PhysiciensDocument7 pagesAlchimie Paracelse (1493-1541) - de La Teinture Des PhysiciensAnri CazoNo ratings yet
- Luxation Traumatique de La HancheDocument4 pagesLuxation Traumatique de La Hanchesahar amerNo ratings yet
- The FaithDocument8 pagesThe FaithGabriel IlungaNo ratings yet
- COURS EN LIGNE DE MIB 4168 Final PDFDocument75 pagesCOURS EN LIGNE DE MIB 4168 Final PDFInnocent Joris Baïbel PouyaNo ratings yet
- Chirurgie GynecologiqueDocument38 pagesChirurgie GynecologiqueLiliana LopesNo ratings yet
- 2 Accidents AdministrationDocument35 pages2 Accidents AdministrationAbd LouahabNo ratings yet
- PRETEST Covid Vaccination Course FRDocument2 pagesPRETEST Covid Vaccination Course FRCrios MNo ratings yet
- Le Nouveau Modèle de Développement Du MarocDocument2 pagesLe Nouveau Modèle de Développement Du MaroctariqNo ratings yet
- Trauma Cranien PDFDocument7 pagesTrauma Cranien PDFyaya camaraNo ratings yet
- Cancers Du FoieDocument60 pagesCancers Du Foieblk fatimaNo ratings yet
- TD2 Denombrement-1Document4 pagesTD2 Denombrement-1Fatima Ezahra RochdiNo ratings yet
- SS Manuel de BiosecuriteDocument42 pagesSS Manuel de BiosecuriteIvy Francine NzouenouNo ratings yet
- Tetu PlusDocument112 pagesTetu PlusMichael MiocheNo ratings yet
- Les Troubles Du Rythme Et de La Conduction CardiaqueDocument53 pagesLes Troubles Du Rythme Et de La Conduction CardiaqueBMA-medecine100% (8)