You might also like
- AcidoDocument6 pagesAcidoDark WolfNo ratings yet
- Pituitary Adenoma-Neuronal Choristoma Is A Pituitary Adenoma WithDocument4 pagesPituitary Adenoma-Neuronal Choristoma Is A Pituitary Adenoma WithRoxana Ioana DumitriuNo ratings yet
- 1 Repiska 2010Document6 pages1 Repiska 2010Thamyres BrancoNo ratings yet
- 2017년 Biomaterials xEntrDocument14 pages2017년 Biomaterials xEntrjonghyeokshin7No ratings yet
- NIH Public Access: Author ManuscriptDocument15 pagesNIH Public Access: Author ManuscriptHutsDMNo ratings yet
- Stem Cell Biology and Regenerative Medicine in OphthalmologyFrom EverandStem Cell Biology and Regenerative Medicine in OphthalmologyStephen TsangNo ratings yet
- 06-cryopreservation of stem cellsDocument29 pages06-cryopreservation of stem cellsjashuj.2004No ratings yet
- Article For DiscussionDocument9 pagesArticle For DiscussionAnthony DavisNo ratings yet
- Trabalho 2 EsteroidaisDocument6 pagesTrabalho 2 EsteroidaisMarcos PecinatoNo ratings yet
- Safitri 2016Document5 pagesSafitri 2016Eng.Químico SCNo ratings yet
- 2011 BakirciogluDocument13 pages2011 BakirciogluMestrado EducacaoNo ratings yet
- Changes in Hippocampal Neurogenesis Throughout Early 2015 Neurobiology of ADocument15 pagesChanges in Hippocampal Neurogenesis Throughout Early 2015 Neurobiology of AErik J. Vallejo FonsecaNo ratings yet
- The National Academies Press: Spinal Cord Injury: Progress, Promise, and Priorities (2005)Document32 pagesThe National Academies Press: Spinal Cord Injury: Progress, Promise, and Priorities (2005)Glenardi HalimNo ratings yet
- Ref 5Document11 pagesRef 5Tiago BaraNo ratings yet
- Divya3 PDFDocument4 pagesDivya3 PDFMekala LakshmanNo ratings yet
- Epididymal Semen Analysis in Testicular Toxicity of Doxorubicin in Male Albino Wistar Rats and Its Amelioration With Quercetin PDFDocument4 pagesEpididymal Semen Analysis in Testicular Toxicity of Doxorubicin in Male Albino Wistar Rats and Its Amelioration With Quercetin PDFMekala LakshmanNo ratings yet
- Apoptosis in Natural Rabies Virus Infection in DogsDocument5 pagesApoptosis in Natural Rabies Virus Infection in DogspapiipiiNo ratings yet
- Inexhaustible Hair-Cell Regeneration in Young and Aged ZebrafishDocument7 pagesInexhaustible Hair-Cell Regeneration in Young and Aged ZebrafishSaifuddin HaswareNo ratings yet
- Pericyte Biology - Novel ConceptsFrom EverandPericyte Biology - Novel ConceptsAlexander BirbrairNo ratings yet
- FullDocument6 pagesFullgerginNo ratings yet
- Referensi Elabscience T2Document12 pagesReferensi Elabscience T2Nandia SeptiyoriniNo ratings yet
- 4nqln1o PDFDocument5 pages4nqln1o PDFGlauce L TrevisanNo ratings yet
- Directing Mouse Embryonic Neurosphere Differentiation Toward An Enriched Neuronal Population - Torrado 2014Document6 pagesDirecting Mouse Embryonic Neurosphere Differentiation Toward An Enriched Neuronal Population - Torrado 2014Rebeca ChávezNo ratings yet
- Advances in Stem Cells and Regenerative Medicine: Single-Cell Dynamics, New Models and Translational PerspectivesDocument5 pagesAdvances in Stem Cells and Regenerative Medicine: Single-Cell Dynamics, New Models and Translational PerspectivesVeridica El EdgieNo ratings yet
- Paper Canty 2009 - PV HolesDocument11 pagesPaper Canty 2009 - PV Holesmaariamunoz9No ratings yet
- Aceto 2015Document42 pagesAceto 2015cepotcopet0101No ratings yet
- Evaluation of The Susceptibility of Neurons Anticancer DrugsDocument6 pagesEvaluation of The Susceptibility of Neurons Anticancer DrugsLisbeth AlmNo ratings yet
- Learn. Mem.-2009-Jessberger-147-54llDocument9 pagesLearn. Mem.-2009-Jessberger-147-54llJean Pierre Chastre LuzaNo ratings yet
- Carbajal BEcellsDocument9 pagesCarbajal BEcellsPatySA2No ratings yet
- A Genetic Mosaic Screen Reveals Ecdysone-Responsive Genes Regulating Drosophila OogenesisDocument15 pagesA Genetic Mosaic Screen Reveals Ecdysone-Responsive Genes Regulating Drosophila Oogenesis奕蓁陳No ratings yet
- Pone 0170735Document13 pagesPone 0170735Vitor PereiraNo ratings yet
- Jneurosci 0073-14 2014Document15 pagesJneurosci 0073-14 2014mahfuz emonNo ratings yet
- Sumber 2Document5 pagesSumber 2Igd An Ni'mahNo ratings yet
- Chao Chuan Wang CV Teaching2013 WDocument6 pagesChao Chuan Wang CV Teaching2013 Wapi-229401491No ratings yet
- 3- 博硕留学生文献阅读2018Document72 pages3- 博硕留学生文献阅读2018fareehakanwar93No ratings yet
- Neural stem cell-derived extracellular vesicles favour neuronal differentiation and plasticity under stress conditionsDocument10 pagesNeural stem cell-derived extracellular vesicles favour neuronal differentiation and plasticity under stress conditionsGedguol AlvarezNo ratings yet
- Xenotransplantation ThesisDocument5 pagesXenotransplantation Thesisafbwrszxd100% (2)
- Ajina Et Al-2017-AndrologiaDocument6 pagesAjina Et Al-2017-AndrologiaMauro Porcel de PeraltaNo ratings yet
- Use of X-Ray Microprobe To Diagnose Bone Tissue Demineralization After Caffeine AdministrationDocument8 pagesUse of X-Ray Microprobe To Diagnose Bone Tissue Demineralization After Caffeine AdministrationbrainNo ratings yet
- Version On-Line ISSN 0717-9502: International Journal of MorphologyDocument8 pagesVersion On-Line ISSN 0717-9502: International Journal of MorphologyMaría AlejandraNo ratings yet
- Prenatal Exposure To Pesticides Disrupts Testicular Histoarchitecture and Alters Testosterone Levels in Male Caiman Latirostris PDFDocument7 pagesPrenatal Exposure To Pesticides Disrupts Testicular Histoarchitecture and Alters Testosterone Levels in Male Caiman Latirostris PDFRiuin23No ratings yet
- Anti-Cancer Potential of Copper Oxide Nanoparticles Against Murine Mammary Adenocarcinoma (Amn-3) CellsDocument6 pagesAnti-Cancer Potential of Copper Oxide Nanoparticles Against Murine Mammary Adenocarcinoma (Amn-3) CellsIJAR JOURNALNo ratings yet
- Exosome Role in Sci RecoveryDocument13 pagesExosome Role in Sci RecoverySilverStarNo ratings yet
- 401 2018 Article 1868Document16 pages401 2018 Article 1868Valen EstevezNo ratings yet
- Tracking in Vitro and in Vivo Sirna Electrotransfer in Tumor CellsDocument8 pagesTracking in Vitro and in Vivo Sirna Electrotransfer in Tumor Cellscindy tatiana rodriguez marinNo ratings yet
- Animals 11 01995Document14 pagesAnimals 11 01995Jihan Natul Sa'diahNo ratings yet
- Clonogenic Assay of Cells in Vitro - Nature Protocol 2006Document5 pagesClonogenic Assay of Cells in Vitro - Nature Protocol 2006Thomas SecherNo ratings yet
- 361 13patilDocument5 pages361 13patilMahi MahaNo ratings yet
- Lou Et Al (2018) - Wound-Healing Effects of 635-nm Low-Level Laser Therapy On Primary Human Vocal Fold Epithelial Cells An in Vitro StudyDocument8 pagesLou Et Al (2018) - Wound-Healing Effects of 635-nm Low-Level Laser Therapy On Primary Human Vocal Fold Epithelial Cells An in Vitro StudyRobson LemosNo ratings yet
- Umbilical Cord Stem Cell Therapy: The Gift of Healing from Healthy NewbornsFrom EverandUmbilical Cord Stem Cell Therapy: The Gift of Healing from Healthy NewbornsNo ratings yet
- Enhancement of direct somatic embryogenesis in Phalaenopsis orchidsDocument8 pagesEnhancement of direct somatic embryogenesis in Phalaenopsis orchidsskullabyNo ratings yet
- CellsDocument14 pagesCellsbibaNo ratings yet
- Organotypic Brain Slices Cultures. 2015Document13 pagesOrganotypic Brain Slices Cultures. 2015Iván LazcanoNo ratings yet
- Hongzhuan Xuan, Zhen Li, Haiyue Yan, Qing Sang, Kai Wang, Qingtao He, Yuanjun WangDocument29 pagesHongzhuan Xuan, Zhen Li, Haiyue Yan, Qing Sang, Kai Wang, Qingtao He, Yuanjun WangTim WongNo ratings yet
- Ref 6Document12 pagesRef 6CARMEN EMILIA RODRIGUEZ SEMINARIONo ratings yet
- Controle Da VentilaçãoDocument7 pagesControle Da VentilaçãoalinestiNo ratings yet
- Deep Learning For Direct Hybrid Precoding in Millimeter Wave Massive MIMO SystemsDocument7 pagesDeep Learning For Direct Hybrid Precoding in Millimeter Wave Massive MIMO SystemspasomagaNo ratings yet
- Array-Based GNSS Signal Tracking With A Reduced State Signal ModelDocument17 pagesArray-Based GNSS Signal Tracking With A Reduced State Signal ModelpasomagaNo ratings yet
- Beyond The Kalman FilterParticle Filters For Tracking ApplicationsDocument47 pagesBeyond The Kalman FilterParticle Filters For Tracking Applicationsnodidino100% (1)
- Encyclopedia of Slavery in The United States PDFDocument910 pagesEncyclopedia of Slavery in The United States PDFpasomagaNo ratings yet
- HW 8Document1 pageHW 8pasomagaNo ratings yet
- Encyclopedia of Slavery in The United States PDFDocument910 pagesEncyclopedia of Slavery in The United States PDFpasomagaNo ratings yet
- L21 Advanced TopicsDocument43 pagesL21 Advanced TopicspasomagaNo ratings yet
- Duchi 16Document88 pagesDuchi 16pasomagaNo ratings yet
- Green Small Cell Operation of Ultra-Dense Networks Using Device AssistanceDocument19 pagesGreen Small Cell Operation of Ultra-Dense Networks Using Device AssistancepasomagaNo ratings yet
- Data-driven baseline estimation for residential demand responseDocument21 pagesData-driven baseline estimation for residential demand responsepasomagaNo ratings yet
- Machine Learning Assignment 1 Basic Concepts: Due: 27 March 2015, 15:00pmDocument3 pagesMachine Learning Assignment 1 Basic Concepts: Due: 27 March 2015, 15:00pmpasomagaNo ratings yet
- 1Document25 pages1pasomagaNo ratings yet
- Complex DiffDocument2 pagesComplex DiffpasomagaNo ratings yet
- Nonparametric Belief Propagation For Self-Localization of Sensor NetworksDocument11 pagesNonparametric Belief Propagation For Self-Localization of Sensor NetworkspasomagaNo ratings yet
- WinEdt Cannot Work With Acrobat Reader 10 (X) Whole TextDocument3 pagesWinEdt Cannot Work With Acrobat Reader 10 (X) Whole TextpasomagaNo ratings yet
- A Distributed Auction Algorithm For The Assignment ProblemDocument6 pagesA Distributed Auction Algorithm For The Assignment ProblempasomagaNo ratings yet
- Energy Systems Integration 101Document86 pagesEnergy Systems Integration 101pasomagaNo ratings yet
- Lecture 1Document28 pagesLecture 1Kamara KumaraNo ratings yet
- Proof of The Cofactor Expansion Theorem 1Document13 pagesProof of The Cofactor Expansion Theorem 1pasomagaNo ratings yet
- Growth, Percolation, and Correlations in Disordered Fiber NetworksDocument29 pagesGrowth, Percolation, and Correlations in Disordered Fiber NetworkspasomagaNo ratings yet
- The MagPi Issue 1 - Raspberry PiDocument32 pagesThe MagPi Issue 1 - Raspberry PiJames BatemanNo ratings yet
- Introduction to Percolation Theory and Critical PhenomenaDocument25 pagesIntroduction to Percolation Theory and Critical Phenomenapasomaga100% (1)
- Balanced Outcomes in Social Exchange NetworksDocument10 pagesBalanced Outcomes in Social Exchange NetworkspasomagaNo ratings yet
- A Cooperation Strategy Based On Nash Bargaining Solution in Cooperative Relay NetworksDocument8 pagesA Cooperation Strategy Based On Nash Bargaining Solution in Cooperative Relay NetworkspasomagaNo ratings yet
- Introduction To Percolation TheoryDocument21 pagesIntroduction To Percolation TheorypasomagaNo ratings yet
- A Survey of Wireless Communication Using Visible LightDocument10 pagesA Survey of Wireless Communication Using Visible LightpasomagaNo ratings yet
- MMC Overview of Multimedia Communication FundamentalsDocument37 pagesMMC Overview of Multimedia Communication FundamentalspasomagaNo ratings yet
- Thesis Olof HerttingDocument57 pagesThesis Olof HerttingAjay IyerNo ratings yet
- E-Techno: Cbse Class-Ix - E6 - E-Techno JEE TEST DATE: 17-11-2020Document16 pagesE-Techno: Cbse Class-Ix - E6 - E-Techno JEE TEST DATE: 17-11-2020Himanshu ThakurNo ratings yet
- TC QMM 56942Document120 pagesTC QMM 56942Fernando R EpilNo ratings yet
- Cosmeceutic ALS: Cosmetics and PharmaceuticalsDocument19 pagesCosmeceutic ALS: Cosmetics and PharmaceuticalssowjanyaNo ratings yet
- DNA TimelineDocument2 pagesDNA TimelineMinaMilićNo ratings yet
- Chapter 5Document14 pagesChapter 5Caitlin G.No ratings yet
- Biology Investigatory ProjectDocument17 pagesBiology Investigatory ProjectAnmol Dhungel100% (6)
- Komiyama Et Al., (2008) Allometry, Biomass, and Productivity of Mangrove Forests A ReviewDocument11 pagesKomiyama Et Al., (2008) Allometry, Biomass, and Productivity of Mangrove Forests A ReviewVandhi Amali100% (1)
- Week 6. Definision and DictionaryDocument19 pagesWeek 6. Definision and DictionaryafdreliautariazizahNo ratings yet
- Lesson 7 - Organ TransplantationDocument14 pagesLesson 7 - Organ TransplantationJosee100% (1)
- 2 Part 1Document99 pages2 Part 1m_luchianNo ratings yet
- General Katalog PT. AmoebaDocument23 pagesGeneral Katalog PT. AmoebaMulyanaNo ratings yet
- Science Holidays Homework Class 9Document2 pagesScience Holidays Homework Class 9nileshNo ratings yet
- Field Inspection Techniques Ifad-CaspDocument17 pagesField Inspection Techniques Ifad-CaspNasiru Kura100% (1)
- Adlerian Psychotherapy: Prioritizing RelationshipsDocument24 pagesAdlerian Psychotherapy: Prioritizing RelationshipsJayanth MamundiNo ratings yet
- A Review of The Environmental Fate, Effects, and Exposures of Bisphenol A PDFDocument25 pagesA Review of The Environmental Fate, Effects, and Exposures of Bisphenol A PDFTiago TorresNo ratings yet
- When The Koel SingsDocument15 pagesWhen The Koel SingsAlfie Paduga BeguinaNo ratings yet
- Explaining Butterfly MetamorphosisDocument13 pagesExplaining Butterfly MetamorphosisCipe drNo ratings yet
- Cardiovascular SystemDocument40 pagesCardiovascular SystemMudasir Hussain TuriNo ratings yet
- Vanders Human Physiology The Mechanisms of Body Function 15th Edition Widmaier Test BankDocument29 pagesVanders Human Physiology The Mechanisms of Body Function 15th Edition Widmaier Test BankDavidWardrcobi100% (15)
- HistamineDocument16 pagesHistaminevg03ik2No ratings yet
- Poultry Science MCQDocument12 pagesPoultry Science MCQelanthamizhmaran83% (6)
- Advt - R-09-2023 WT ResultDocument16 pagesAdvt - R-09-2023 WT ResultAvinash Kumar SinghNo ratings yet
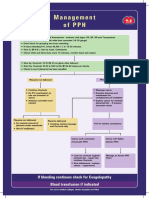
- Management of PPHDocument1 pageManagement of PPH098 U.KARTHIK SARAVANA KANTHNo ratings yet
- 10th Standard Tamilnadu State Board Science (English Medium)Document283 pages10th Standard Tamilnadu State Board Science (English Medium)Karthick NNo ratings yet
- C Specialized GeneralBiology1 Sem1 Clas3 CellsinPlantand-AnimalTissue v2Document14 pagesC Specialized GeneralBiology1 Sem1 Clas3 CellsinPlantand-AnimalTissue v2ダギア メイ アニアNo ratings yet
- The "Five Families" College Essay ExampleDocument1 pageThe "Five Families" College Essay ExampleKishor RaiNo ratings yet
- Pain PathwayDocument17 pagesPain PathwaySalsabila Al-BasheerNo ratings yet
- Artigo Sobre Nutrigenômica e Nutrigenética PDFDocument18 pagesArtigo Sobre Nutrigenômica e Nutrigenética PDFDiogo JuniorNo ratings yet
- Gazzaniga The Ethical - Brain.the - Science.of - Our.moral - DilemmasDocument229 pagesGazzaniga The Ethical - Brain.the - Science.of - Our.moral - Dilemmaskid_latigo100% (9)