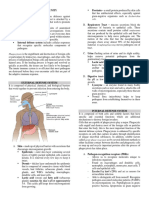
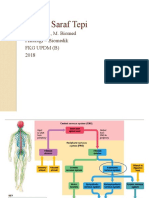
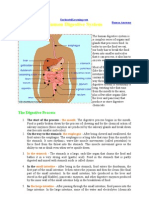
You might also like
- Immunology Janis Kuby .Page001Document1 pageImmunology Janis Kuby .Page001microkannanNo ratings yet
- A Gut Feeling For Immune Dysregulation & Neuroinflammation in Autism by Aristo Vojdani, PHD, MSC, MT and Jama LambertDocument9 pagesA Gut Feeling For Immune Dysregulation & Neuroinflammation in Autism by Aristo Vojdani, PHD, MSC, MT and Jama LambertautismoneNo ratings yet
- Inflammatory and Immunologic DisturbancesDocument91 pagesInflammatory and Immunologic DisturbancesRitamaria100% (6)
- 7 - Regulation and Functions of The Thyroid HormonesDocument31 pages7 - Regulation and Functions of The Thyroid HormonesVigneshwaran RavishankarNo ratings yet
- CH1 Path D&R AgamDocument34 pagesCH1 Path D&R Agam062100% (1)
- ImmunologyDocument36 pagesImmunologyمروه عماد عيسى100% (1)
- Immunology KubyDocument24 pagesImmunology KubySayanta Bera100% (1)
- 1 - Anatomy of The KidneyDocument15 pages1 - Anatomy of The Kidneysara khanNo ratings yet
- Lecture 2.3 - Defense MechanismsDocument40 pagesLecture 2.3 - Defense MechanismsNiña Viaña BinayNo ratings yet
- Lesson 10 Brain Parts, Processes and FunctionsDocument19 pagesLesson 10 Brain Parts, Processes and FunctionsAlfredo ModestoNo ratings yet
- Infectious Disorders HandoutsDocument35 pagesInfectious Disorders Handoutsfebie pachecoNo ratings yet
- EmbryologyDocument68 pagesEmbryologyBj Larracas100% (1)
- Human ReproductionDocument20 pagesHuman ReproductionKhibul Lim100% (1)
- Autoimmune Disease MechanismsDocument6 pagesAutoimmune Disease MechanismsHaliunaa BattulgaNo ratings yet
- The Slim Book of Health Pearls: Man the Barricades - The Story of the Immune SystemFrom EverandThe Slim Book of Health Pearls: Man the Barricades - The Story of the Immune SystemNo ratings yet
- Review Flash Card by Dr. Mitch Roy MaristelaDocument205 pagesReview Flash Card by Dr. Mitch Roy MaristelaMark Justin OcampoNo ratings yet
- Testicular Torsion: Dr. Hari Krismanuel, Sp. B, FinacsDocument19 pagesTesticular Torsion: Dr. Hari Krismanuel, Sp. B, FinacsVanessa Modi AlverinaNo ratings yet
- Ce Sunt InfectiileDocument5 pagesCe Sunt Infectiilegeorgi.annaNo ratings yet
- Non Specific and Specific Host Defense MechanismDocument7 pagesNon Specific and Specific Host Defense Mechanismcheskhadomingo33No ratings yet
- VMC 221 Coursematerials PDFDocument61 pagesVMC 221 Coursematerials PDFavenger 2100% (1)
- Chapter 33 Prescott Innate Immunity PDFDocument7 pagesChapter 33 Prescott Innate Immunity PDFneeru.bhagatNo ratings yet
- Immunology & Serology ReviewerDocument71 pagesImmunology & Serology ReviewerMark Justin OcampoNo ratings yet
- MIcro - Immunology - Lec.1Document13 pagesMIcro - Immunology - Lec.1زين العابدين محمد عويش مشريNo ratings yet
- Immunopharmacology - IntroductionDocument51 pagesImmunopharmacology - IntroductionIsrat Jahan Surovy 2225400672No ratings yet
- NCM 112 Midterm Notes 2Document67 pagesNCM 112 Midterm Notes 2ljealou2002No ratings yet
- Chapter 16Document5 pagesChapter 16Alanah JaneNo ratings yet
- FinaDocument19 pagesFinaZabdiel Ann SavellanoNo ratings yet
- Sworts Immunology - Introduction-To-Part-IDocument8 pagesSworts Immunology - Introduction-To-Part-IꕶᗋᏦᎥꕗNo ratings yet
- IMMUNITYDocument4 pagesIMMUNITYSoni VishwakarmaNo ratings yet
- Prelims - GMJ Lecture - Module 2 III DisorderDocument5 pagesPrelims - GMJ Lecture - Module 2 III DisorderjuiceNo ratings yet
- The Immune System: Lindsay B. NicholsonDocument27 pagesThe Immune System: Lindsay B. NicholsonAlanakimiNo ratings yet
- NCM 112: Care of Clients With Inflammatory and Immunologic Response Maricris B. Florendo/Constantino Y. Mendoza/Sandra Layne G. Luczon/Rhizza AbinsayDocument23 pagesNCM 112: Care of Clients With Inflammatory and Immunologic Response Maricris B. Florendo/Constantino Y. Mendoza/Sandra Layne G. Luczon/Rhizza AbinsayRyrey Abraham PacamanaNo ratings yet
- Chapter 1 - MicrobioDocument5 pagesChapter 1 - MicrobioFaith ManiquisNo ratings yet
- Bio PoemDocument3 pagesBio Poemapi-4039173890% (1)
- Immune System HandoutDocument7 pagesImmune System HandoutRaineir PabiranNo ratings yet
- BB Week 2Document7 pagesBB Week 2Claudille GawaranNo ratings yet
- The Immune System ExplainedDocument2 pagesThe Immune System Explainedvilmar montejoNo ratings yet
- Lec # 4 Body Defence SystemsDocument31 pagesLec # 4 Body Defence SystemssamotherlianNo ratings yet
- Introduction To Immune SystemDocument37 pagesIntroduction To Immune SystemPatrick MukosoNo ratings yet
- Full Immunity MaterialDocument93 pagesFull Immunity MaterialKajetano GraucosNo ratings yet
- Immune System ProjectDocument21 pagesImmune System ProjectAvinash BobadeNo ratings yet
- IMMUNITY - (Allied Health Sciences) - Complete - March 2024Document51 pagesIMMUNITY - (Allied Health Sciences) - Complete - March 2024Anania EmmanuelNo ratings yet
- Body Defenses & Immunity: But If Skin Is Moist, Not Cleaned Frequently EnoughDocument10 pagesBody Defenses & Immunity: But If Skin Is Moist, Not Cleaned Frequently EnoughSherwann Vergara Delos ReyesNo ratings yet
- BCM 127 Immunology Lecture Series 2020 01Document77 pagesBCM 127 Immunology Lecture Series 2020 01Barnabas KipngetichNo ratings yet
- Body Defences, Immunity, ImmunizationDocument27 pagesBody Defences, Immunity, Immunizationenam professorNo ratings yet
- Innate Immunity: OutlineDocument8 pagesInnate Immunity: OutlineJane BacnotanNo ratings yet
- Vaccine Immunology: Understanding Modern Vaccines: Perspectives in VaccinologyDocument35 pagesVaccine Immunology: Understanding Modern Vaccines: Perspectives in VaccinologyNabila AzzaetunaNo ratings yet
- Sas Hes032 9Document6 pagesSas Hes032 9Jose Melmar Autida AutenticoNo ratings yet
- CH 15 Intro To Immune ResponseDocument4 pagesCH 15 Intro To Immune Responseericka abasNo ratings yet
- In Ammation, Immunity and Allergy: Learning ObjectivesDocument6 pagesIn Ammation, Immunity and Allergy: Learning ObjectivesJavier VeraNo ratings yet
- Innate Immunity Chapter SummaryDocument11 pagesInnate Immunity Chapter SummaryOrhan AsdfghjklNo ratings yet
- NCM 119 - Rle (Prelim) Week 1: Immune Response Immunity: Natural and Acquired Immunity - Ma'am VAB Immune ResponseDocument13 pagesNCM 119 - Rle (Prelim) Week 1: Immune Response Immunity: Natural and Acquired Immunity - Ma'am VAB Immune ResponseJamaica Leslie Noveno100% (1)
- Immunity 190910064848 PDFDocument78 pagesImmunity 190910064848 PDFDeepak KumarNo ratings yet
- DSM Immunity WhitepaperDocument18 pagesDSM Immunity WhitepaperSilambarasu KaruppiahNo ratings yet
- Fundamental of VaccineDocument7 pagesFundamental of VaccinecarlsonrenovatioNo ratings yet
- Immune SystemDocument56 pagesImmune SystemGerome CausapinNo ratings yet
- An Introduction To Immunology and Immunopathology: Allergy, Asthma & Clinical ImmunologyDocument10 pagesAn Introduction To Immunology and Immunopathology: Allergy, Asthma & Clinical Immunologyhendra cahyaditaNo ratings yet
- Aisyah Umayi - Infografis PatologiDocument1 pageAisyah Umayi - Infografis PatologirarasNo ratings yet
- IMMUNOLOGY - 3 - Innate ImmunityDocument5 pagesIMMUNOLOGY - 3 - Innate ImmunityFelix NepumocenoNo ratings yet
- Chapter 15 - Innate ImmunityDocument7 pagesChapter 15 - Innate ImmunityAlicia Marie ElizaldeNo ratings yet
- Understanding the Animal Immune SystemDocument2 pagesUnderstanding the Animal Immune SystemDan Jonathan De VegaNo ratings yet
- Lecture 7. CH - 16 - Lecture - Presentation Innate Immunity 2019 (NJ)Document68 pagesLecture 7. CH - 16 - Lecture - Presentation Innate Immunity 2019 (NJ)Moshe Cohen'sNo ratings yet
- Physical Activities, Exercise and Their Effects To Inmune SystemDocument7 pagesPhysical Activities, Exercise and Their Effects To Inmune SystemCarolina Bejarano GonzalezNo ratings yet
- K Upper 2004Document12 pagesK Upper 2004Võ Thành DuyNo ratings yet
- Lec 1Document24 pagesLec 1Ab AbNo ratings yet
- Imse Trans PrelimDocument12 pagesImse Trans PrelimSarah EugenioNo ratings yet
- Control and Prevention of Communicable DiseasesDocument14 pagesControl and Prevention of Communicable DiseasesZabdiel Ann SavellanoNo ratings yet
- Immunology Unveiled: A Comprehensive Journey through the Human Immune System: Guardians of the Body: The Unseen Heroes of ImmunityFrom EverandImmunology Unveiled: A Comprehensive Journey through the Human Immune System: Guardians of the Body: The Unseen Heroes of ImmunityNo ratings yet
- SciGenom Trace Viewer Control Sample AnalysisDocument1 pageSciGenom Trace Viewer Control Sample AnalysisVigneshwaran RavishankarNo ratings yet
- Faculty Advt 6000AGPDocument21 pagesFaculty Advt 6000AGPSri DNo ratings yet
- Additions IIDocument3 pagesAdditions IISakthidasan SankaranNo ratings yet
- Perfect M.EDocument28 pagesPerfect M.EVigneshwaran RavishankarNo ratings yet
- The Six Sigma Principle PDFDocument7 pagesThe Six Sigma Principle PDFVigneshwaran RavishankarNo ratings yet
- LeptospirosisDocument4 pagesLeptospirosisasril111No ratings yet
- Taguchis Quality Loss FunctionDocument6 pagesTaguchis Quality Loss FunctionVigneshwaran RavishankarNo ratings yet
- EBT108Document1 pageEBT108Vigneshwaran RavishankarNo ratings yet
- LeptospirosisDocument4 pagesLeptospirosisasril111No ratings yet
- BiochemDocument1 pageBiochemLiezyl Dela CruzNo ratings yet
- Biotech 2013Document11 pagesBiotech 2013Vigneshwaran RavishankarNo ratings yet
- A Quantitative PCR (Taqman) Assay For Pathogenic: Leptospira SPPDocument22 pagesA Quantitative PCR (Taqman) Assay For Pathogenic: Leptospira SPPVigneshwaran RavishankarNo ratings yet
- A Quantitative PCR (Taqman) Assay For Pathogenic: Leptospira SPPDocument22 pagesA Quantitative PCR (Taqman) Assay For Pathogenic: Leptospira SPPVigneshwaran RavishankarNo ratings yet
- J. Clin. Microbiol.-1992-Mérien-2219-24Document7 pagesJ. Clin. Microbiol.-1992-Mérien-2219-24Vigneshwaran RavishankarNo ratings yet
- Family JesusDocument3 pagesFamily JesusVigneshwaran RavishankarNo ratings yet
- Supporting InfoDocument4 pagesSupporting InfoVigneshwaran RavishankarNo ratings yet
- CTSDocument5 pagesCTSVigneshwaran RavishankarNo ratings yet
- Upload FilesDocument1 pageUpload FilesVigneshwaran RavishankarNo ratings yet
- NDA TemplateDocument3 pagesNDA TemplateVigneshwaran RavishankarNo ratings yet
- NDA TemplateDocument3 pagesNDA TemplateVigneshwaran RavishankarNo ratings yet
- What Is AIDS?: Acquired Immunodeficiency Syndrome (AIDS) Is A Collection of Diseases ResultingDocument5 pagesWhat Is AIDS?: Acquired Immunodeficiency Syndrome (AIDS) Is A Collection of Diseases ResultingVigneshwaran RavishankarNo ratings yet
- NoteDocument1 pageNoteVigneshwaran RavishankarNo ratings yet
- Enzyme and Microbial TechnologyDocument8 pagesEnzyme and Microbial TechnologyVigneshwaran RavishankarNo ratings yet
- Unit Test II Questions on Biopharmaceutical TechnologyDocument2 pagesUnit Test II Questions on Biopharmaceutical TechnologyVigneshwaran RavishankarNo ratings yet
- Suggested Answers To Exercise, Reading To Learn and Cross-Topic ExerciseDocument19 pagesSuggested Answers To Exercise, Reading To Learn and Cross-Topic ExerciseBernardNo ratings yet
- 8.2 # Human Digestive SystemDocument5 pages8.2 # Human Digestive SystemSara Nadeem KhanNo ratings yet
- DentinDocument89 pagesDentinDr Zadeno kithanNo ratings yet
- AneurysmDocument11 pagesAneurysmNashrah HusnaNo ratings yet
- RADIOLOGY ASPECT OF EXTRA PULMONARY TUBERCULOSISDocument45 pagesRADIOLOGY ASPECT OF EXTRA PULMONARY TUBERCULOSISAndre F SusantioNo ratings yet
- Abdominal TraumaDocument30 pagesAbdominal TraumaKavya KesavanNo ratings yet
- Animal Tissues WorksheetDocument8 pagesAnimal Tissues WorksheetMaxene LuisNo ratings yet
- Fisiologi Sistem Saraf TepiDocument19 pagesFisiologi Sistem Saraf TepiNata NayottamaNo ratings yet
- Acute Biologic Crisis QuizDocument1 pageAcute Biologic Crisis Quiznurse_maeNo ratings yet
- Autoimmune Hemolytic Anemia in Children - Classification, Clinical Features, and Diagnosis - UpToDateDocument20 pagesAutoimmune Hemolytic Anemia in Children - Classification, Clinical Features, and Diagnosis - UpToDateNedelcu Miruna100% (1)
- Cutaneous CirculationDocument19 pagesCutaneous CirculationShubhamNo ratings yet
- Science 9, LM 2020Document427 pagesScience 9, LM 2020Mira VeranoNo ratings yet
- Lesson Plan for Hematology-IVDocument4 pagesLesson Plan for Hematology-IVRenuka KhatkarNo ratings yet
- Male Reproductive System CanvaDocument35 pagesMale Reproductive System CanvaDex VargasNo ratings yet
- Basic Concepts of Hematology (Hemostasis)Document53 pagesBasic Concepts of Hematology (Hemostasis)Hendra SshNo ratings yet
- Self CareDocument27 pagesSelf CareJessa May RapadasNo ratings yet
- All in One Golden CPSP QuestionsDocument570 pagesAll in One Golden CPSP QuestionsQuranSunnat100% (2)
- VahimetsmartinDocument0 pagesVahimetsmartinAngela PagliusoNo ratings yet
- Pathology: HematologyDocument1 pagePathology: HematologyMubashar AliNo ratings yet
- SEM 6 - 10 - BSC - HONS - MICROBIOLOGY - CC-13 - IMMUNOLOGY - 10109Document3 pagesSEM 6 - 10 - BSC - HONS - MICROBIOLOGY - CC-13 - IMMUNOLOGY - 10109soumyadip shitNo ratings yet
- PHC 513 Flipped Class QuestionDocument17 pagesPHC 513 Flipped Class QuestionALISYA SOPHIA MOHAMMAD ABU SHAHID CHRISNo ratings yet
- Digestive System Label MeDocument13 pagesDigestive System Label MeMuhammad AdibNo ratings yet
- Adult Hippocampal NeurogenesisDocument14 pagesAdult Hippocampal NeurogenesisElaine HermansNo ratings yet