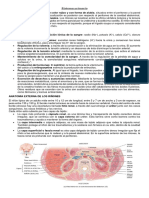
You might also like
- Fisiología Renal: C.D Yarlim Lima YarmaDocument15 pagesFisiología Renal: C.D Yarlim Lima YarmaAshh LuNo ratings yet
- Aparato Urinario Marco TeòricoDocument18 pagesAparato Urinario Marco TeòricoAlxAgorNo ratings yet
- Guia Renal DefinitivaDocument8 pagesGuia Renal DefinitivaDouglas Soslan Reyes FiffeNo ratings yet
- Sistema UrinarioDocument12 pagesSistema UrinarioosmitsalazarNo ratings yet
- RiñónDocument14 pagesRiñónJose Alberto Lazcano BrunoNo ratings yet
- Sistema UrinarioDocument5 pagesSistema UrinarioPaula ElvazNo ratings yet
- Sistema Urinario Histologia Uabp 1Document7 pagesSistema Urinario Histologia Uabp 1lua robertiNo ratings yet
- 11 Aparato Urinario - Apunte de CátedraDocument9 pages11 Aparato Urinario - Apunte de CátedraLuz OlivaNo ratings yet
- Manual de exploración urinariaDocument80 pagesManual de exploración urinariacampsar50% (2)
- Aparato UrinarioDocument19 pagesAparato UrinarioAnaNo ratings yet
- Histologia - Resumen de Aparato UrinarioDocument11 pagesHistologia - Resumen de Aparato UrinarioValeria MéndezNo ratings yet
- Powers Renal UnidosDocument210 pagesPowers Renal UnidosPaula ProezaNo ratings yet
- Histologia Sistema UrinarioDocument4 pagesHistologia Sistema Urinariocheveloso100% (2)
- Anatomía Macroscópica y Microscópica Del RiñónDocument7 pagesAnatomía Macroscópica y Microscópica Del RiñónKarina Del Valle0% (1)
- La Nefrona, Tubulos Proximales y DistalesDocument2 pagesLa Nefrona, Tubulos Proximales y DistalesHeidi Esther BaezNo ratings yet
- Gartner, Leslie P. (19 Sistema Urinario)Document24 pagesGartner, Leslie P. (19 Sistema Urinario)Ardila NubiaNo ratings yet
- Histología Sistema UrinarioDocument8 pagesHistología Sistema UrinarioEngerber PerezNo ratings yet
- Sistema excretor: riñones y nefronasDocument7 pagesSistema excretor: riñones y nefronasRonny Pita Murillo100% (1)
- Resumen Sistema UrinarioDocument6 pagesResumen Sistema UrinariogabrielledeluqNo ratings yet
- Anatomía y Fisiología RenalDocument10 pagesAnatomía y Fisiología RenalJoan william Salazar paguasiNo ratings yet
- Anatomía y Fisiología Del Sistema UrinarioDocument7 pagesAnatomía y Fisiología Del Sistema UrinarioMarcos LeonNo ratings yet
- Sistema Urinario EN PDFDocument7 pagesSistema Urinario EN PDFDa CostaNo ratings yet
- Aparato Urinario y Sistema ReproductorDocument14 pagesAparato Urinario y Sistema ReproductorEnos DEGBENo ratings yet
- Anatomia y Fisiologia Renal Del Riñon - ResumenDocument8 pagesAnatomia y Fisiologia Renal Del Riñon - ResumenEstephany MolinaNo ratings yet
- Función y estructura del riñónDocument3 pagesFunción y estructura del riñónDaniel Alberto Garcia AguilarNo ratings yet
- Catedra de NefrologiaDocument41 pagesCatedra de NefrologiaMarcus MunozNo ratings yet
- Sistema UrinarioDocument35 pagesSistema Urinariohenriquezrodriguez2023No ratings yet
- Definición Del Sistema UrinarioDocument8 pagesDefinición Del Sistema UrinarioMichael Joseph EsquivelNo ratings yet
- Apuntes RenalDocument3 pagesApuntes RenalLuis JuarezNo ratings yet
- ¿Que Es El Riñon?Document4 pages¿Que Es El Riñon?Vero PerezNo ratings yet
- Configuración interna del riñón: corteza, médula y lóbulos renalesDocument6 pagesConfiguración interna del riñón: corteza, médula y lóbulos renalesVanessa ManríquezNo ratings yet
- Anatomía Del RiñónDocument4 pagesAnatomía Del RiñónMaite PashmayNo ratings yet
- Cs. Biológicas - Sist. RenalDocument59 pagesCs. Biológicas - Sist. RenalAna belen GomezNo ratings yet
- guia renalDocument9 pagesguia renalFrank MontezaNo ratings yet
- Fisiologia RenalDocument16 pagesFisiologia Renalrafo18No ratings yet
- Dinamica AnatoDocument7 pagesDinamica AnatoANGEL23No ratings yet
- Anatomia y Fisiologia Del RiñonDocument70 pagesAnatomia y Fisiologia Del RiñonNorberto GarciaNo ratings yet
- Aparato UrinarioDocument64 pagesAparato UrinarioSalvador BARRON MELONo ratings yet
- Tarea Del Primer Parcial de GenitourinarioDocument9 pagesTarea Del Primer Parcial de GenitourinarioRetdiniNo ratings yet
- Tema 8 ArregloDocument30 pagesTema 8 ArregloGonzalo Sánchez DiéguezNo ratings yet
- Aparato UrinarioDocument11 pagesAparato UrinarioDariiana Benavides100% (1)
- Sistema urinario: riñones, nefronas y función renalDocument7 pagesSistema urinario: riñones, nefronas y función renalemil obeidNo ratings yet
- Gartner, Leslie P. - Texto Atlas de Histologia, 2da Edición (19 Sistema Urinario) PDFDocument24 pagesGartner, Leslie P. - Texto Atlas de Histologia, 2da Edición (19 Sistema Urinario) PDFAarón E. Carías100% (1)
- Sistema urinario: estructura y función de los riñonesDocument20 pagesSistema urinario: estructura y función de los riñonesPatricia PetersNo ratings yet
- SISTEMA UrinarioDocument27 pagesSISTEMA UrinarioJuan Carlos DOrso100% (1)
- NefronaDocument8 pagesNefronaluisestradai1310No ratings yet
- RiñonesDocument4 pagesRiñoneskanousitaNo ratings yet
- RIÑÓNDocument7 pagesRIÑÓNParker GreenNo ratings yet
- Resumen Sistema UrinarioDocument9 pagesResumen Sistema UrinarioMarina Morales100% (1)
- histologia riñones puntosDocument13 pageshistologia riñones puntosgabriela escobarNo ratings yet
- AnatomíaDocument66 pagesAnatomíaMarìa Daniela ZambranoNo ratings yet
- Tejido MuscularDocument9 pagesTejido MuscularDaniela HeLoNo ratings yet
- Histología Del RiñonDocument6 pagesHistología Del RiñonKarime LopezavilaNo ratings yet
- Apuntes Sistema Urinario TM 4Document11 pagesApuntes Sistema Urinario TM 4Isamí C.No ratings yet
- Fisiopatologia RenalDocument82 pagesFisiopatologia RenalAbraham Bejarano CoronelNo ratings yet
- Cartilla Aparato urinario Unidad 9Document19 pagesCartilla Aparato urinario Unidad 9Lucas RuizNo ratings yet
- Fisiologia RenalDocument40 pagesFisiologia RenalNoryeski LopezNo ratings yet
- Coagulación Para Todos: Medicina Para TodosFrom EverandCoagulación Para Todos: Medicina Para TodosRating: 4.5 out of 5 stars4.5/5 (3)
- Introducción Básica a la Microcirculación de la Sangre: La Siguiente Generación en Cuidados de la SaludFrom EverandIntroducción Básica a la Microcirculación de la Sangre: La Siguiente Generación en Cuidados de la SaludNo ratings yet
- Anemia Para Todos: Medicina Para TodosFrom EverandAnemia Para Todos: Medicina Para TodosRating: 3 out of 5 stars3/5 (1)
- Ingredientes Exóticos: Propiedades y Aplicaciones DermofarmacéuticasDocument8 pagesIngredientes Exóticos: Propiedades y Aplicaciones DermofarmacéuticasAndres RestrepoNo ratings yet
- 1922-Texto Del Artículo-6337-1-10-20190204Document7 pages1922-Texto Del Artículo-6337-1-10-20190204Eliana Maria Sierra JimenezNo ratings yet
- Clase 2 - PneDocument23 pagesClase 2 - PneAndres RestrepoNo ratings yet
- Índice de Función Sexual FemeninaDocument4 pagesÍndice de Función Sexual FemeninaAndres RestrepoNo ratings yet
- El DesdichadoDocument5 pagesEl DesdichadoAndres RestrepoNo ratings yet
- Presentación DiverticulitisDocument17 pagesPresentación DiverticulitisAndres RestrepoNo ratings yet
- Trabajo de Recuperación NordesteDocument2 pagesTrabajo de Recuperación NordesteAndres RestrepoNo ratings yet
- (4131) Diciembre 29 de 2015 Publicado 30 de Diciembre de 2015Document358 pages(4131) Diciembre 29 de 2015 Publicado 30 de Diciembre de 2015Andres RestrepoNo ratings yet
- Orgasmo y CerebroDocument5 pagesOrgasmo y CerebroAndres RestrepoNo ratings yet
- Clase 1 - PneDocument23 pagesClase 1 - PneAndres RestrepoNo ratings yet
- Trabajo de Recuperación NordesteDocument2 pagesTrabajo de Recuperación NordesteAndres RestrepoNo ratings yet
- Sexualidad & Diversidad FuncionalDocument109 pagesSexualidad & Diversidad FuncionalGeorge Viera100% (2)
- Sca Con Elevación Del STDocument20 pagesSca Con Elevación Del STAndres Restrepo100% (1)
- El DesdichadoDocument5 pagesEl DesdichadoAndres RestrepoNo ratings yet
- Presentación AsmaDocument20 pagesPresentación AsmaAndres RestrepoNo ratings yet
- Guías para Manejo de Urgencias Tomo IiiDocument600 pagesGuías para Manejo de Urgencias Tomo IiiAngelik M Hernández100% (11)
- Guías para Manejo de Urgencias Tomo IiDocument632 pagesGuías para Manejo de Urgencias Tomo IiDavid Fernando Lozada Poveda100% (4)
- Definicion de EnfermeriaDocument1 pageDefinicion de EnfermeriaAndres RestrepoNo ratings yet
- Guias para Manejo de Urgencias Tomo IDocument624 pagesGuias para Manejo de Urgencias Tomo IDavid Fernando Lozada Poveda83% (6)
- Lexico Griego-Español NT. Alfred TuggyDocument1,163 pagesLexico Griego-Español NT. Alfred TuggyJosePerez100% (16)
- El Divorcio Es Permitido para Que La Maldad No Llegue A Proporciones InsosteniblesDocument17 pagesEl Divorcio Es Permitido para Que La Maldad No Llegue A Proporciones InsosteniblesAndres RestrepoNo ratings yet
- Estudio Transfer Factor Muy BuenoDocument29 pagesEstudio Transfer Factor Muy BuenoAndres RestrepoNo ratings yet
- Guias para Manejo de Urgencias Tomo IDocument624 pagesGuias para Manejo de Urgencias Tomo IDavid Fernando Lozada Poveda83% (6)
- Curso Ministerial Apóstol Juan BallistreriDocument1 pageCurso Ministerial Apóstol Juan BallistreriAndres RestrepoNo ratings yet
- Descristianizando El CristianismoDocument2 pagesDescristianizando El CristianismoAndres RestrepoNo ratings yet
- Bosquejo Pablo HechosDocument4 pagesBosquejo Pablo HechosAndres RestrepoNo ratings yet
- Triage PrehospitalarioDocument53 pagesTriage PrehospitalarioAndres RestrepoNo ratings yet
- Estructura de Ahorro 2018Document8 pagesEstructura de Ahorro 2018Andres RestrepoNo ratings yet
- PEI CENCALA Colombia VERSION 02 08 DE SEPTIEMBRE DE 2016 PDFDocument64 pagesPEI CENCALA Colombia VERSION 02 08 DE SEPTIEMBRE DE 2016 PDFarman373No ratings yet
- Guias para Manejo de Urgencias Tomo IDocument624 pagesGuias para Manejo de Urgencias Tomo IDavid Fernando Lozada Poveda83% (6)