You might also like
- Grupos de Insectos Según Sus Habitos de AlimentaciónDocument12 pagesGrupos de Insectos Según Sus Habitos de AlimentaciónCarlos Herrera100% (1)
- 01 Historia de La MicologíaDocument11 pages01 Historia de La MicologíaJesus Bautisto Huichi Jara100% (1)
- Montaje de InsectosDocument9 pagesMontaje de InsectosMiriam RodríguezNo ratings yet
- DípterosDocument12 pagesDípterosCarlaLagosMoralesNo ratings yet
- Clostridium Corynebacterium MycobacteriumDocument66 pagesClostridium Corynebacterium MycobacteriumMiguel Alva SevillaNo ratings yet
- Introducción Hongos 3Document3 pagesIntroducción Hongos 3Fernanda Torres RojasNo ratings yet
- Técnica de Necropsia de RumiantesDocument86 pagesTécnica de Necropsia de RumiantesTotoNo ratings yet
- Apuntes Parasitologia LDocument154 pagesApuntes Parasitologia LBrenda Alexandra Cardenas BedoyaNo ratings yet
- Patogenos de Plantas Descritos en EspañaDocument858 pagesPatogenos de Plantas Descritos en EspañaZhirley Mariley100% (1)
- Preservación y Montaje de InsectosDocument3 pagesPreservación y Montaje de InsectosFrancisco GarcíaNo ratings yet
- Quorum SensingDocument2 pagesQuorum SensingCoraima GuevaraNo ratings yet
- Microcultivo ArticuloDocument3 pagesMicrocultivo ArticuloMartín Daniel ParraNo ratings yet
- Tema 3 Eucarya HongosDocument75 pagesTema 3 Eucarya Hongoslady maritza parra orjuelaNo ratings yet
- Historia de La Fitopatologia en Ica (Aproximacion)Document9 pagesHistoria de La Fitopatologia en Ica (Aproximacion)Christian Eduardo Quispe TorresNo ratings yet
- Genoma Eucariota Modificado Final PDFDocument33 pagesGenoma Eucariota Modificado Final PDFCarmen BarbozaNo ratings yet
- Presentacion Unidad II MICOLOGÍADocument89 pagesPresentacion Unidad II MICOLOGÍASalim PavonNo ratings yet
- Miasis PDFDocument33 pagesMiasis PDFMia Obando Velasquez100% (1)
- Album de Parasitos - Jazury Olea MoralesDocument29 pagesAlbum de Parasitos - Jazury Olea MoralesJazzie OM 대No ratings yet
- Trabajo Tarantula 26-9-17Document14 pagesTrabajo Tarantula 26-9-17JonathanAguilarNo ratings yet
- VESPIDAEDocument9 pagesVESPIDAELorena Toro100% (1)
- AcantocefalosDocument52 pagesAcantocefalosCarlos D. Luján SifuentesNo ratings yet
- Aplicador de PlaguicidasDocument400 pagesAplicador de Plaguicidascarlos_j_nNo ratings yet
- Sanidad Vegetal Hongos JandryDocument6 pagesSanidad Vegetal Hongos JandryJandry ZambranoNo ratings yet
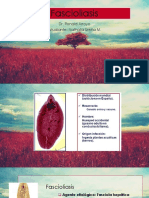
- Fascioliasis 3 ExamenDocument30 pagesFascioliasis 3 ExamenNatyure XzNo ratings yet
- ARTRÓPODOS DE INTERÉS MÉDICO EN ARGENTINA Mundo Sano MonograDocument112 pagesARTRÓPODOS DE INTERÉS MÉDICO EN ARGENTINA Mundo Sano MonograVanesa MelgarezNo ratings yet
- Kit Básico Guia Cultivo Fungico Mantun Grow.Document8 pagesKit Básico Guia Cultivo Fungico Mantun Grow.NonsensesNo ratings yet
- Proyecto para Cultivo de Acelga en Proceso Por TerminarDocument15 pagesProyecto para Cultivo de Acelga en Proceso Por Terminarmiltonq97100% (1)
- JORGE1Document54 pagesJORGE1Jorge Luis Flores RojasNo ratings yet
- Hongos: Características, clasificación y reproducciónDocument45 pagesHongos: Características, clasificación y reproducciónjacson ortuñoNo ratings yet
- Glosario BotánicaDocument37 pagesGlosario BotánicaDiego Morodo67% (3)
- Insectos ChupadoresDocument24 pagesInsectos ChupadoresandresNo ratings yet
- Cuestionario de Esterilización y Desinfección 1Document2 pagesCuestionario de Esterilización y Desinfección 1ismael fernandez zarateNo ratings yet
- PRÁCTICA 3 Aislamiento-CultivoDocument16 pagesPRÁCTICA 3 Aislamiento-CultivoLuis Angel Azañero RumayNo ratings yet
- Reconocimiento de síntomas y signos en plantasDocument7 pagesReconocimiento de síntomas y signos en plantasfernandorodriguez284gmail.comNo ratings yet
- Asepsia y BioseguridadDocument1 pageAsepsia y BioseguridadHayna Casas CastroNo ratings yet
- Trips y Hormiga Antenas - Aparato BucalDocument3 pagesTrips y Hormiga Antenas - Aparato BucalaldairNo ratings yet
- Preparación de abono orgánico BocashiDocument8 pagesPreparación de abono orgánico BocashiAB KHEYLA XIMENA RIVAS MARTINEZNo ratings yet
- Observación de estructuras fúngicasDocument7 pagesObservación de estructuras fúngicasArlinson LlanosNo ratings yet
- 01 Exposición Granos Almacenados NNDocument74 pages01 Exposición Granos Almacenados NNYURY EDSON CHAVEZ100% (1)
- Silvestres Comestibles de Setas Y Trufas de OregonDocument2 pagesSilvestres Comestibles de Setas Y Trufas de Oregonquestionablescr61No ratings yet
- Fiebre AftosaDocument14 pagesFiebre AftosaAngie YaimaNo ratings yet
- Guía TerminadaDocument46 pagesGuía TerminadaJohnny Saavedra CamachoNo ratings yet
- Control de Plagas y Enfermedades de Las PlantasDocument81 pagesControl de Plagas y Enfermedades de Las PlantasVerdeamorNo ratings yet
- Estructuras Fúngicas de Hongos de Interés Clínico y AmbientalDocument12 pagesEstructuras Fúngicas de Hongos de Interés Clínico y AmbientalFredy RuizNo ratings yet
- Clasificacion de HongosDocument15 pagesClasificacion de HongosLupita OlanNo ratings yet
- Historia de La FitopatologiaDocument2 pagesHistoria de La FitopatologiaOKBZASNo ratings yet
- Campylobacteriosis: causas, síntomas y prevenciónDocument19 pagesCampylobacteriosis: causas, síntomas y prevenciónjhon molinaNo ratings yet
- Clase 4 Taxonomia de Hongos Biodiversidad IDocument69 pagesClase 4 Taxonomia de Hongos Biodiversidad IAnonymous dplPP0iNo ratings yet
- NutriDocument15 pagesNutrisamyucraNo ratings yet
- Muestreo FitopatológicoDocument12 pagesMuestreo FitopatológicoLuis FernandoNo ratings yet
- NEMATODODocument32 pagesNEMATODOJosmell GamboaNo ratings yet
- Micología ClínicaDocument8 pagesMicología ClínicaJatniel DelgadoNo ratings yet
- Introducción a los artrópodos de interés médicoDocument49 pagesIntroducción a los artrópodos de interés médicoJesusDanielHernandezEscheikNo ratings yet
- 4-Phylum Myxomycota 10Document6 pages4-Phylum Myxomycota 10Ybeth GalvezNo ratings yet
- 17 Himenolepiasis..Document23 pages17 Himenolepiasis..Rafael CintiaNo ratings yet
- Diapositivas de Mohos y LevadurasDocument25 pagesDiapositivas de Mohos y LevadurasWendyRamosNo ratings yet
- Hongos Resumen y Apuntes SellDocument10 pagesHongos Resumen y Apuntes SellAnahi López100% (1)
- TECNICA DE COLECTA Y PRESERVACION DE INSECTOS TEMA 1 Y 2jDocument18 pagesTECNICA DE COLECTA Y PRESERVACION DE INSECTOS TEMA 1 Y 2jKarina VargasNo ratings yet
- Hongos Nematofagos PDFDocument3 pagesHongos Nematofagos PDFJorge TerzoNo ratings yet
- Expo 2 Nematodo 1Document30 pagesExpo 2 Nematodo 1Milagros EspinozaNo ratings yet
- Tif - PiñaDocument12 pagesTif - PiñaGladisMidaRodriguezLastraNo ratings yet
- Practica 6 PlatanosDocument7 pagesPractica 6 PlatanosGladisMidaRodriguezLastraNo ratings yet
- Control ExpoDocument12 pagesControl ExpoGladisMidaRodriguezLastraNo ratings yet
- 4.-Monografia de InjertosDocument13 pages4.-Monografia de InjertosGladisMidaRodriguezLastra83% (6)
- 4.-Monografia de InjertosDocument13 pages4.-Monografia de InjertosGladisMidaRodriguezLastra83% (6)
- Instalacion ParabolicaDocument28 pagesInstalacion Parabolicapolifonico100% (2)
- Crianza y Produccion de Conservas de TruchaDocument33 pagesCrianza y Produccion de Conservas de TruchaCANALES VIRU, Marlon Ivan100% (1)
- Cons Trucci OnDocument12 pagesCons Trucci OnKaren Carreon QuezadaNo ratings yet
- Informe Pascua LamaDocument39 pagesInforme Pascua LamaBioBioChileNo ratings yet
- Es 05625 La-De Emt PDFDocument73 pagesEs 05625 La-De Emt PDFINGJDIAZNo ratings yet
- Ime 076Document177 pagesIme 076ivanortega16RNo ratings yet
- Estudio MOrtero - Cemeto.ceniza - Rastrojo.maizDocument12 pagesEstudio MOrtero - Cemeto.ceniza - Rastrojo.maizWilder Rimac MendezNo ratings yet
- ENCOFRADOSDocument36 pagesENCOFRADOSLeyser Elí Cerdan ChuquimangoNo ratings yet
- Procesos Tecnológicos para La Obtención Del AceroDocument13 pagesProcesos Tecnológicos para La Obtención Del AceroDonaldo Jzpd100% (2)
- EVALUANDO PropiedadesDocument2 pagesEVALUANDO Propiedadesmaggi79No ratings yet
- Bradley Cat 07Document12 pagesBradley Cat 07FERCONo ratings yet
- HIDROMETALURGIADocument9 pagesHIDROMETALURGIAJerson Ramos HuertaNo ratings yet
- Pliego Especificaciones Eca FinalDocument18 pagesPliego Especificaciones Eca FinalMarisa AlarconNo ratings yet
- 1partes de Un Mci - MandarDocument26 pages1partes de Un Mci - MandarQuinta LunaNo ratings yet
- Manual de Procedimientos para Manejo de Madera Proveniente Del ProyectoDocument25 pagesManual de Procedimientos para Manejo de Madera Proveniente Del ProyectounanuevasabanaNo ratings yet
- Ciencia e Ingeniería de Materiales-Conceptos de Metalurgia FísicaDocument13 pagesCiencia e Ingeniería de Materiales-Conceptos de Metalurgia FísicaDanielAlamoNo ratings yet
- Eficiencia ElectromecanicaDocument12 pagesEficiencia ElectromecanicaJavier Ruiz O100% (1)
- Tomate PDFDocument8 pagesTomate PDFNacho Garcia ThorNo ratings yet
- Formación integral y orientación laboralDocument40 pagesFormación integral y orientación laboralPerezNo ratings yet
- NMX-L-140-1995 AtapulguitaDocument16 pagesNMX-L-140-1995 AtapulguitaFernando García RamónNo ratings yet
- Informe de Ensayo de Corte DirectoDocument14 pagesInforme de Ensayo de Corte DirectoJaner GilNo ratings yet
- Tithonia DiversifoliaDocument6 pagesTithonia DiversifoliaMax Lenin EstradaNo ratings yet
- Normas AASHTO LRFD para diseño y características de puentesDocument5 pagesNormas AASHTO LRFD para diseño y características de puentesCHRISTHIAN RICARDO GARCÍA REYESNo ratings yet
- Validation of The Gold Determination MetDocument16 pagesValidation of The Gold Determination MetAndrés PacompíaNo ratings yet
- Catálogo de Productos Industrias SaladilloDocument33 pagesCatálogo de Productos Industrias SaladilloXimena May100% (3)
- Producción artesanal y industrial de jabónDocument73 pagesProducción artesanal y industrial de jabónmilagros bailon sanchezNo ratings yet
- Elaboración de cuchillaDocument12 pagesElaboración de cuchillamarthaisabelroa2817100% (2)
- Diseño y construcción de un calentador solar de agua por termosifónDocument12 pagesDiseño y construcción de un calentador solar de agua por termosifónCesar JeanNo ratings yet
- DebercompletoDocument15 pagesDebercompletovicky m0% (1)
- 440-05 Código de Colores P' Equipos de S-EDocument16 pages440-05 Código de Colores P' Equipos de S-EJuan Santiago Bolivar GomezNo ratings yet