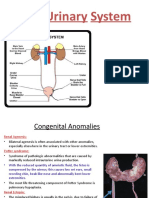
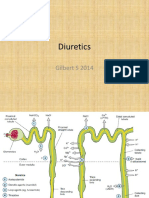
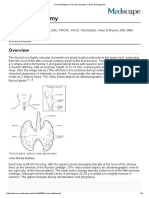
You might also like
- Kidney Functions, Anatomy and ProcessesDocument48 pagesKidney Functions, Anatomy and ProcessesMsatriaNo ratings yet
- Lecture Notes On The Renal System-1Document51 pagesLecture Notes On The Renal System-1GiftedNo ratings yet
- ExcertionDocument39 pagesExcertionMuhammad UsmanNo ratings yet
- HTC2-Physiology: Structure and Function of KidneyDocument47 pagesHTC2-Physiology: Structure and Function of KidneyVienne Yuen Wing YanNo ratings yet
- Renal Anatomy and Physiology: SrsibalDocument49 pagesRenal Anatomy and Physiology: SrsibalAbraham DemeterioNo ratings yet
- Urinary System: by Eka BebasariDocument47 pagesUrinary System: by Eka BebasariMuhammad Reza SaputraNo ratings yet
- Renal Physiology: by Dr. Isam Eldin Mohamed Abd AllaDocument34 pagesRenal Physiology: by Dr. Isam Eldin Mohamed Abd AllaSiddigNo ratings yet
- Filtrasi GlomerulusDocument41 pagesFiltrasi GlomerulusFaishalNo ratings yet
- Excretory SystemDocument55 pagesExcretory SystemBhargavi PriyaNo ratings yet
- Urinary SystemDocument84 pagesUrinary SystemCheghe PerezNo ratings yet
- Renal System 2022Document78 pagesRenal System 2022Rahil PatelNo ratings yet
- The Urinary System: DR P VijayalakshmiDocument79 pagesThe Urinary System: DR P VijayalakshmiVijaya LakshmiNo ratings yet
- 2-Renal Physiology 2 (Renal Haemodynamic and GFR)Document28 pages2-Renal Physiology 2 (Renal Haemodynamic and GFR)Iph anyiNo ratings yet
- Chapter 15 Student Version The Urinary System 2020.ppt (1402)Document20 pagesChapter 15 Student Version The Urinary System 2020.ppt (1402)S. MartinezNo ratings yet
- Urine Formation. PDF My NoteDocument11 pagesUrine Formation. PDF My NoteSurya JoyNo ratings yet
- Urine FormationDocument4 pagesUrine FormationSoumya RathNo ratings yet
- Urinary System: G R O U P 4 - B S N 1 - 5Document59 pagesUrinary System: G R O U P 4 - B S N 1 - 5Love Jovellanos ÜNo ratings yet
- The Urinary SystemDocument76 pagesThe Urinary SystemDeborah Chimeremeze JosephNo ratings yet
- Mechanism of Urine FormationDocument35 pagesMechanism of Urine FormationHemanth PrakashNo ratings yet
- 1 RENAL Glomerular Nephrotic2016Document45 pages1 RENAL Glomerular Nephrotic2016Crystal Lynn Keener SciariniNo ratings yet
- Vertebrate Kidney Functions and ProcessesDocument57 pagesVertebrate Kidney Functions and Processesnokate konkoorNo ratings yet
- Multinucleated (Many Nuclei) : Osmoregulation: Is The Control of The Water Balance of The Blood, Tissue or Cytoplasm of ADocument2 pagesMultinucleated (Many Nuclei) : Osmoregulation: Is The Control of The Water Balance of The Blood, Tissue or Cytoplasm of Atae gukNo ratings yet
- Notes For HomeostasisDocument21 pagesNotes For Homeostasisfreednemon80% (5)
- Determinants of Glomerular Filtration RateDocument52 pagesDeterminants of Glomerular Filtration RateAbraham Dawson machachaNo ratings yet
- Renal PathophysiologyDocument19 pagesRenal PathophysiologyMcKinley SwiftNo ratings yet
- Anatomy and Physiology of the Urinary System ExplainedDocument53 pagesAnatomy and Physiology of the Urinary System ExplainedArathy KalathingalNo ratings yet
- Topic 2b - Homeostasis-Student v2Document52 pagesTopic 2b - Homeostasis-Student v2Angel LimNo ratings yet
- Lecture 11 - Urinary SystemDocument64 pagesLecture 11 - Urinary SystemmaecabauatanNo ratings yet
- The Urinary System: William J. Babler, PH.DDocument30 pagesThe Urinary System: William J. Babler, PH.DMary-GraceScalesNo ratings yet
- Urinary SystemDocument68 pagesUrinary SystemWilson chumpukaNo ratings yet
- Renal Physiology 2: Urine Formation and Regulation of GFRDocument39 pagesRenal Physiology 2: Urine Formation and Regulation of GFRHema KamatNo ratings yet
- Filtrasi GlomerulusDocument26 pagesFiltrasi GlomerulusA1205Angelica GloryNo ratings yet
- Urinary System-1Document86 pagesUrinary System-1greatmoosaNo ratings yet
- CH 26 UrinaryDocument63 pagesCH 26 Urinaryaijaz1inNo ratings yet
- Kidney 1Document41 pagesKidney 1Miguel Cuevas DolotNo ratings yet
- PhysiologyDocument85 pagesPhysiologyDyah WahliaNo ratings yet
- Clinical Anatomy of Kidney: NephronDocument3 pagesClinical Anatomy of Kidney: NephronNeil AlviarNo ratings yet
- Anatomy and Physiology of The Urinary System: Dipali HarkhaniDocument43 pagesAnatomy and Physiology of The Urinary System: Dipali HarkhaniZala KarmitNo ratings yet
- Kidney PhysiologyDocument144 pagesKidney PhysiologySilasNo ratings yet
- Histology of the Urinary SystemDocument48 pagesHistology of the Urinary SystemNel TinduganiNo ratings yet
- 5 6249228362582068405Document63 pages5 6249228362582068405Isha NagleNo ratings yet
- Kidney Anatomy: NephronDocument3 pagesKidney Anatomy: NephronNeil AlviarNo ratings yet
- Excretory Products AND Their EliminationDocument43 pagesExcretory Products AND Their EliminationBiju MylachalNo ratings yet
- Anatomy and Physiology of The KidneysDocument21 pagesAnatomy and Physiology of The Kidneysccbrown750% (2)
- Function of KidneyDocument66 pagesFunction of KidneyNilesh100% (4)
- Glomerular Filtration Rate RegulationDocument13 pagesGlomerular Filtration Rate RegulationMarilia BonorinoNo ratings yet
- Fluid Compartments, Intracellular CompartmentsVinoDocument28 pagesFluid Compartments, Intracellular CompartmentsVinongidizzmNo ratings yet
- Anatomy & Physiology of The Urinary System.: Presented By:-Mr. Saihou Sanneh RN, RM, BSN&RH, FwacnDocument70 pagesAnatomy & Physiology of The Urinary System.: Presented By:-Mr. Saihou Sanneh RN, RM, BSN&RH, Fwacnflex gyNo ratings yet
- Structure and Function of Kidneys: HTC2-Physiology-26-10-21 DR CH LaiDocument4 pagesStructure and Function of Kidneys: HTC2-Physiology-26-10-21 DR CH LaiVienne Yuen Wing YanNo ratings yet
- Urinary System and Nephron AnatomyDocument59 pagesUrinary System and Nephron AnatomyLove Jovellanos ÜNo ratings yet
- Seminar MicrocirculaionDocument48 pagesSeminar MicrocirculaionKirtishAcharyaNo ratings yet
- Histology of The Urinary System: BY DR C.M. OkekeDocument62 pagesHistology of The Urinary System: BY DR C.M. OkekeonyekaNo ratings yet
- What Are AntibioticsDocument29 pagesWhat Are AntibioticsLouie BruanNo ratings yet
- Kidney Function Lecture NotesDocument80 pagesKidney Function Lecture NotesNomel JonesNo ratings yet
- Group 10 Urine FormationDocument27 pagesGroup 10 Urine FormationgracechamdimbaNo ratings yet
- Excretory Products and Their EliminationDocument9 pagesExcretory Products and Their EliminationBiju MylachalNo ratings yet
- LESSON 3 Renal Function and TestsDocument63 pagesLESSON 3 Renal Function and TestsFaith TambongNo ratings yet
- Histology of Urinary System F - 084046Document69 pagesHistology of Urinary System F - 084046Malak NourNo ratings yet
- Kidney Diseases, A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsFrom EverandKidney Diseases, A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsNo ratings yet
- Symposium: Cardiac Arrhythmias (Part 2)Document10 pagesSymposium: Cardiac Arrhythmias (Part 2)Josephine SNo ratings yet
- Effects and mechanisms of diuretics in the kidneyDocument34 pagesEffects and mechanisms of diuretics in the kidneyJosephine SNo ratings yet
- Kidney 2013 P4P DiscussionDocument17 pagesKidney 2013 P4P DiscussionJosephine SNo ratings yet
- Hydrocephalus PathophysiologyDocument2 pagesHydrocephalus PathophysiologyJosephine SNo ratings yet
- Skin Physiology: Lecturer:Dr Herman Mulijadi MS, SPKPDocument59 pagesSkin Physiology: Lecturer:Dr Herman Mulijadi MS, SPKPJosephine SNo ratings yet
- Anat Week 5Document3 pagesAnat Week 5Josephine SNo ratings yet
- 12 Chapter 3Document29 pages12 Chapter 3Josephine SNo ratings yet
- 3 QaDocument5 pages3 QaJosephine SNo ratings yet
- Everything You Need to Know About Hair and NailsDocument2 pagesEverything You Need to Know About Hair and NailsJosephine SNo ratings yet
- Thyroid Anatomy - Overview, Structure, Fascia and LigamentDocument6 pagesThyroid Anatomy - Overview, Structure, Fascia and LigamentJosephine SNo ratings yet
- Manual of Lab. Endocrine Week 4 2017Document3 pagesManual of Lab. Endocrine Week 4 2017Josephine SNo ratings yet
- Fluids and Electrolytes NCM 101Document142 pagesFluids and Electrolytes NCM 101France John Evangelista Torres100% (2)
- 17 - Toronto Notes 2011 - NephrologyDocument92 pages17 - Toronto Notes 2011 - NephrologyKhairulsani Yusof100% (2)
- Renal SystemDocument14 pagesRenal SystemFrances RebeccaNo ratings yet
- Urine Formation: Mohamad Ashraf IsmailDocument13 pagesUrine Formation: Mohamad Ashraf IsmailAshraf MobyNo ratings yet
- A&P Target ScoresDocument72 pagesA&P Target ScoresMercurie8592% (12)
- Ondansetron: Gastric EmptyingDocument4,170 pagesOndansetron: Gastric EmptyingdrsaidumerNo ratings yet
- Urinalysis and Body Fluids 6th Edition by Susan King Strasinger - Test BankDocument45 pagesUrinalysis and Body Fluids 6th Edition by Susan King Strasinger - Test Bankroseyoung0No ratings yet
- Pcol 211 Prelims ALL inDocument413 pagesPcol 211 Prelims ALL inPaula LouiseNo ratings yet
- Daily Practice Test-14 (Zoology) Questions: Excretory SystemDocument7 pagesDaily Practice Test-14 (Zoology) Questions: Excretory SystemDarshan DarshuNo ratings yet
- Renal Function TestsDocument10 pagesRenal Function TestsVirendraNo ratings yet
- Fluid and Electrolyte TherapyDocument23 pagesFluid and Electrolyte TherapyJamal ArizonaNo ratings yet
- Drug Use in Special Conditions - CLD and CKDDocument93 pagesDrug Use in Special Conditions - CLD and CKDadamu mohammadNo ratings yet
- Dr. Naitik D Trivedi & Dr. Upama N. Trivedi: Multiple Choice Questions (Analysis and Chemistry)Document35 pagesDr. Naitik D Trivedi & Dr. Upama N. Trivedi: Multiple Choice Questions (Analysis and Chemistry)Seema YadavNo ratings yet
- Care of The Patient With Renal and Urinary Disorders - HandoutDocument52 pagesCare of The Patient With Renal and Urinary Disorders - HandoutAaron Ramirez LuNo ratings yet
- Renal & Urinary Systems Exam Study GuideDocument9 pagesRenal & Urinary Systems Exam Study GuideAllison GajadharNo ratings yet
- Kidney HealthDocument77 pagesKidney Healthksenthil77No ratings yet
- Allied Exam Answer Key Errors Matter To CM by Vijay Kumar HeerDocument19 pagesAllied Exam Answer Key Errors Matter To CM by Vijay Kumar HeerVIJAY KUMAR HEERNo ratings yet
- Aubf Review UermDocument18 pagesAubf Review UermZhiddah Japzon0% (1)
- Aubf Lec and LabDocument214 pagesAubf Lec and LabAubrey TablateNo ratings yet
- IGCSE Excretion in Humans (Edited)Document59 pagesIGCSE Excretion in Humans (Edited)elizabethNo ratings yet
- Chronic Kidney Disease Case PresDocument32 pagesChronic Kidney Disease Case Presnnaesor_1091No ratings yet
- Amerman Sample Chapter24Document44 pagesAmerman Sample Chapter24Rajan VermaNo ratings yet
- PT Gi and Renal ExamDocument10 pagesPT Gi and Renal ExamNnaa LlppNo ratings yet
- Excretory System PresentationDocument12 pagesExcretory System Presentationapi-263389150No ratings yet
- Egfr ChartDocument1 pageEgfr ChartMahesh DoshiNo ratings yet
- Fluid, Electrolyte and Acid-Base BalanceDocument74 pagesFluid, Electrolyte and Acid-Base BalanceIgwe SolomonNo ratings yet
- Gaddam, Ikshwak: ResultDocument1 pageGaddam, Ikshwak: Resultpraveen kumarNo ratings yet
- Renal Physiology Acid Base BalanceDocument136 pagesRenal Physiology Acid Base BalanceTaimoor Ul HassanNo ratings yet
- Excretory System ResonanceDocument40 pagesExcretory System ResonanceEkta ManglaniNo ratings yet
- Excrection 1Document18 pagesExcrection 1Liyana AziziNo ratings yet