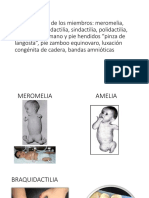
You might also like
- Llavero Tablas de MultiplicarDocument10 pagesLlavero Tablas de MultiplicarGuillermo May100% (1)
- Tipos de inmunidad: Innata, adaptativa y sus componentesDocument29 pagesTipos de inmunidad: Innata, adaptativa y sus componentes05-EF-HU-ELOYZA URIETH GOMEZ PONCENo ratings yet
- WHODAS 2.0 Auto AdministrableDocument5 pagesWHODAS 2.0 Auto AdministrableCcarlos JuannNo ratings yet
- Cancer - Herencia y Ambiente - Cristina CortinasDocument50 pagesCancer - Herencia y Ambiente - Cristina Cortinascarabe100% (3)
- Guía para La Elaboración de Monografía Como Opción A TitulaciónDocument53 pagesGuía para La Elaboración de Monografía Como Opción A TitulaciónJulissa Ruiz CamposNo ratings yet
- InmunologiaDocument45 pagesInmunologiaElizabeth ArévaloNo ratings yet
- Simulacro Examen Oposiciones Técnico Analisis ClinicosDocument38 pagesSimulacro Examen Oposiciones Técnico Analisis ClinicosJesica PerezNo ratings yet
- RQC y SRQDocument5 pagesRQC y SRQAlejandro ArboledaNo ratings yet
- II Unidad Morfofisiologia III Completa Samuel ReyesDocument252 pagesII Unidad Morfofisiologia III Completa Samuel ReyesDiego HernandezNo ratings yet
- Ejercicios de lectura para bachillerato sobre nomofobiaDocument44 pagesEjercicios de lectura para bachillerato sobre nomofobiaMax Mar100% (4)
- EL SISTEMA INMUNITARIO - Diapositivas 2° EtapaDocument33 pagesEL SISTEMA INMUNITARIO - Diapositivas 2° Etapacosas del coleNo ratings yet
- Inmunología Celular y MolecularDocument6 pagesInmunología Celular y MolecularJose VelasquezNo ratings yet
- Inmunidad innata: barreras y mecanismos de defensa naturalDocument5 pagesInmunidad innata: barreras y mecanismos de defensa naturalKarla TorresNo ratings yet
- Inmunidad y AlergiaDocument13 pagesInmunidad y AlergiaAlejandra100% (1)
- Año Del Fortalecimiento de La Soberania Naciona1Document7 pagesAño Del Fortalecimiento de La Soberania Naciona1Analu AndradeNo ratings yet
- Procesos Fisiológicos, El Sistema Inmunitario 2Document23 pagesProcesos Fisiológicos, El Sistema Inmunitario 2Laura Montes Raya100% (1)
- 7 AÑO - CIENCIAS NATURALES - 2021 - SISTEMA INMUNE - CLASE 3 y 4Document5 pages7 AÑO - CIENCIAS NATURALES - 2021 - SISTEMA INMUNE - CLASE 3 y 4Ignacio CatalánNo ratings yet
- Documento 99Document11 pagesDocumento 99Angeliannis CardenadNo ratings yet
- Inmunologia Medica PDFDocument106 pagesInmunologia Medica PDFMariaCieloPfNo ratings yet
- InmunofisiologíaDocument37 pagesInmunofisiologíaLuchitoGoicocheaTrauco100% (1)
- Respuesta InmunitariaDocument33 pagesRespuesta InmunitariaDani UrdaNo ratings yet
- Mecanismo de ResistenciaDocument14 pagesMecanismo de ResistenciaAnonymous 7MbQoo5oQNo ratings yet
- Introducción al sistema inmunitario: células, inmunidades innata y adaptativaDocument10 pagesIntroducción al sistema inmunitario: células, inmunidades innata y adaptativaandreaNo ratings yet
- Primera Clase InmunologiaDocument47 pagesPrimera Clase InmunologiaIvonne Mercedes Gutierrez AtapaucarNo ratings yet
- Inmuno ProyectoDocument27 pagesInmuno ProyectoMichelle YañezNo ratings yet
- Sistema InmuneDocument15 pagesSistema InmuneViviana Carrion AbollanedaNo ratings yet
- Tema 1 - Introducción INMUNOLOGIADocument134 pagesTema 1 - Introducción INMUNOLOGIAHugo Martínez MarínNo ratings yet
- Inmunología Microbiana: Resistencia y AnticuerposDocument44 pagesInmunología Microbiana: Resistencia y AnticuerposJavier SevillaNo ratings yet
- Wuolah-Free-Inmunologia Completo PDFDocument105 pagesWuolah-Free-Inmunologia Completo PDFanon_580890214100% (1)
- Apuntes de InmunoDocument103 pagesApuntes de InmunoCarolinaNo ratings yet
- Apuntes de InmunologiaDocument127 pagesApuntes de InmunologiaLeon Alcon100% (2)
- Respuesta Inmmune MicrobiologiaDocument7 pagesRespuesta Inmmune MicrobiologiaAndrea Gissel Mart�nez BocanegraNo ratings yet
- Universidad Tecnológica de SantiagoDocument12 pagesUniversidad Tecnológica de SantiagoAmdres AlmonteNo ratings yet
- Anexo N - 1 Sistema InmunitarioDocument5 pagesAnexo N - 1 Sistema InmunitarioRocio ColmanNo ratings yet
- Inmunología ApuntesDocument12 pagesInmunología ApuntesclauNo ratings yet
- Guía N°2 Octavo Tercer PeriodoDocument11 pagesGuía N°2 Octavo Tercer PeriodoLuis Miguel Lopez EspinosaNo ratings yet
- Unidad 11-Inmunidad y Organos LinfaticosDocument11 pagesUnidad 11-Inmunidad y Organos LinfaticosYaz OchoaNo ratings yet
- Vacunas 7 MosDocument3 pagesVacunas 7 MosBárbara OyarzoNo ratings yet
- Inmunologia PreguntasDocument6 pagesInmunologia PreguntasIVONNE GERALDINE SANTACRUZ AMADORNo ratings yet
- Texto Base Inmunología - Parte 1Document140 pagesTexto Base Inmunología - Parte 1milamilianchavezNo ratings yet
- SUEROS Y VACUNAS Sistema InmunitarioDocument10 pagesSUEROS Y VACUNAS Sistema InmunitarioMary JiménezNo ratings yet
- InmunidadDocument7 pagesInmunidadMelanie Monserrat Sosa ArteagaNo ratings yet
- Actividades Fisiopatología InmunologíaDocument4 pagesActividades Fisiopatología InmunologíaInes Retamar MartelNo ratings yet
- InmunidadDocument12 pagesInmunidadLulu PerezNo ratings yet
- Lección 1Document42 pagesLección 1Fabiola Rojas JiménezNo ratings yet
- Capitulo 1 InmunologíaDocument6 pagesCapitulo 1 InmunologíamartinvillarrealuasmedicinaNo ratings yet
- Sistema inmune: células, barreras y respuestasDocument3 pagesSistema inmune: células, barreras y respuestasRonald De La CruzNo ratings yet
- SISTEMA INMUNE TrabajoDocument26 pagesSISTEMA INMUNE Trabajomaria sotoNo ratings yet
- Guia InmunologíaDocument10 pagesGuia InmunologíaFatima JTNo ratings yet
- Capítulo 34 GuytonDocument12 pagesCapítulo 34 GuytonCecilia VásquezNo ratings yet
- INMUNOLOGÍA Y SISTEMA INMUNEDocument54 pagesINMUNOLOGÍA Y SISTEMA INMUNEAna JimenezNo ratings yet
- Conceptos Basicos de InmunologiaDocument3 pagesConceptos Basicos de InmunologiaYohanly Hernández AlmonteNo ratings yet
- La InmunidadDocument3 pagesLa InmunidadMari HONo ratings yet
- 2DA - BIOLOGIA.g2.. InmunidadDocument3 pages2DA - BIOLOGIA.g2.. InmunidadAlex Salazar DávilaNo ratings yet
- Tipos de Inmunidad Clase 2 MicrobiologiaDocument4 pagesTipos de Inmunidad Clase 2 MicrobiologiaMarko RamOsNo ratings yet
- Bases Moleculares de La InmunidadDocument24 pagesBases Moleculares de La InmunidadLUIS MATEO PARAMO SANTANDERNo ratings yet
- T5 Respuesta InmuneDocument11 pagesT5 Respuesta InmunearturofernandezgraciaNo ratings yet
- Tema Numero 2 de Medicina TropicalDocument31 pagesTema Numero 2 de Medicina TropicalELBA ROJAS CLAROSNo ratings yet
- 2022 Salesiano. Sistema Inmunitario.Document4 pages2022 Salesiano. Sistema Inmunitario.Alejandro GimenezNo ratings yet
- Tarea 1 Semana 2-GUIADocument4 pagesTarea 1 Semana 2-GUIAJosefina JiménezNo ratings yet
- Inmunología - Generalidades. (1) UmDocument12 pagesInmunología - Generalidades. (1) UmFlorencia RamirezNo ratings yet
- Apuntes FYTI (2009)Document154 pagesApuntes FYTI (2009)Cristina Coronilla BatresNo ratings yet
- Sistema InmuneDocument5 pagesSistema InmunePatricio andres Villarroel GonzalezNo ratings yet
- Inmunidad innata y adquirida: diferencias claveDocument6 pagesInmunidad innata y adquirida: diferencias claveMARCOS ADRIANO FERREIRA DAVIDNo ratings yet
- Defensas Del OrganismoDocument3 pagesDefensas Del OrganismoJOSE MANUEL SALDAA AGUALIMPIANo ratings yet
- Resumen InmunoDocument5 pagesResumen InmunoAmayrani BustamanteNo ratings yet
- Respuesta Inmune (Inmunologia)Document14 pagesRespuesta Inmune (Inmunologia)Joyce JuarezNo ratings yet
- Cuál Es El Concepto de Inmunidad Cuando Se Esta Iniciando La InmunologiaDocument10 pagesCuál Es El Concepto de Inmunidad Cuando Se Esta Iniciando La InmunologiaFlavio MontezaNo ratings yet
- EurekaDocument12 pagesEurekaFlavio MontezaNo ratings yet
- EurekaDocument12 pagesEurekaFlavio MontezaNo ratings yet
- Imagenes de CuestionarioDocument7 pagesImagenes de CuestionarioFlavio MontezaNo ratings yet
- Clase 01 (02-03) CMADocument45 pagesClase 01 (02-03) CMAFlavio MontezaNo ratings yet
- Causalidad y Diagnostico de Enfermedad Ocup. y AccidDocument58 pagesCausalidad y Diagnostico de Enfermedad Ocup. y AccidVane BustamanteNo ratings yet
- Listeria-Campy-Helicobacter PDFDocument20 pagesListeria-Campy-Helicobacter PDFFlavio MontezaNo ratings yet
- Pentosas ClaseDocument15 pagesPentosas ClaseFlavio MontezaNo ratings yet
- Sistema RespiratorioDocument2 pagesSistema RespiratorioFlavio MontezaNo ratings yet
- Tema 8 PresentacionDocument32 pagesTema 8 PresentacionFlavio MontezaNo ratings yet
- Anemia MegaloblasticaDocument13 pagesAnemia MegaloblasticaFlavio MontezaNo ratings yet
- 03 Intolerancia A La Lactosa Art11 PDFDocument11 pages03 Intolerancia A La Lactosa Art11 PDFFlavio MontezaNo ratings yet
- Desarrollo Del Sistema Osteo Muscular MalformacionesDocument33 pagesDesarrollo Del Sistema Osteo Muscular MalformacionesFlavio MontezaNo ratings yet
- 4.conceptos Teoricos PDFDocument73 pages4.conceptos Teoricos PDFpercy riofrio ninoNo ratings yet
- Base Xxvii Eureka 2017 PDFDocument38 pagesBase Xxvii Eureka 2017 PDFVictor Hugo Cardenas LozanoNo ratings yet
- Desarrollo Del Sistema Osteo Muscular MalformacionesDocument3 pagesDesarrollo Del Sistema Osteo Muscular MalformacionesFlavio MontezaNo ratings yet
- Sistema EsqueleticoDocument15 pagesSistema EsqueleticoyusegelmarinoNo ratings yet
- ESPERMATOGÉNESISDocument6 pagesESPERMATOGÉNESISFlavio MontezaNo ratings yet
- Pregunta 5 EmbrioDocument4 pagesPregunta 5 EmbrioFlavio MontezaNo ratings yet
- Pierna 2018Document54 pagesPierna 2018Flavio MontezaNo ratings yet
- Placenta MicroscópicADocument12 pagesPlacenta MicroscópicAFlavio MontezaNo ratings yet
- Estructura 1 Fisiología de La Membrana CelularDocument1 pageEstructura 1 Fisiología de La Membrana CelularFlavio MontezaNo ratings yet
- 09 17 Aminas Heterociclos 17 Chi HeliDocument39 pages09 17 Aminas Heterociclos 17 Chi HelilualmunzuNo ratings yet
- Pregunta 5 EmbrioDocument10 pagesPregunta 5 EmbrioFlavio MontezaNo ratings yet
- ESPERMATOGÉNESISDocument6 pagesESPERMATOGÉNESISFlavio MontezaNo ratings yet
- Los ValoresDocument12 pagesLos ValoresJuan Manuel Reina Montoya100% (1)
- Desarrollo Del Sistema Osteo Muscular MalformacionesDocument33 pagesDesarrollo Del Sistema Osteo Muscular MalformacionesFlavio MontezaNo ratings yet
- 09 17 Aminas Heterociclos 17 Chi HeliDocument39 pages09 17 Aminas Heterociclos 17 Chi HelilualmunzuNo ratings yet
- 09 17 Aminas Heterociclos 17 Chi HeliDocument39 pages09 17 Aminas Heterociclos 17 Chi HeliFlavio MontezaNo ratings yet
- Aparato de GolgiDocument2 pagesAparato de Golgijuan david castillo bernalNo ratings yet
- Informe de La VaricelaDocument4 pagesInforme de La VaricelaJose RoseroNo ratings yet
- Guía de TPs NeurofisiologíaDocument80 pagesGuía de TPs NeurofisiologíaIgnacio VarelaNo ratings yet
- Modulo 3 Prodelab ProhibidoDocument56 pagesModulo 3 Prodelab ProhibidoIsabel Peña MaldonadoNo ratings yet
- Fisiologia Digestiva y Nutricion en La Especie CunicolaDocument21 pagesFisiologia Digestiva y Nutricion en La Especie CunicolaMaclaux SanchezNo ratings yet
- Análisis de Vida de Ellen Basada enDocument7 pagesAnálisis de Vida de Ellen Basada enpedroNo ratings yet
- Tomografía Por Emisión de Positrones: Universidad Privada Del ValleDocument36 pagesTomografía Por Emisión de Positrones: Universidad Privada Del ValleMatheus Santana CedoNo ratings yet
- Lectura s8Document10 pagesLectura s8LUIS EDSON PAUL CRUZ DAVILANo ratings yet
- Taller 1. Historia Epidemiologia 2023 - 2Document3 pagesTaller 1. Historia Epidemiologia 2023 - 2ADRIANA ERASONo ratings yet
- Cultivo in Vitro de FresaDocument25 pagesCultivo in Vitro de FresaDeyler Iglesias100% (1)
- Caso Clinico para RossanaDocument42 pagesCaso Clinico para RossanazoravickNo ratings yet
- 6.presentación Funciones Exponenciales y LogarítmicasDocument18 pages6.presentación Funciones Exponenciales y LogarítmicasLeslie SanchezNo ratings yet
- Delirium TremensDocument2 pagesDelirium TremensyaaqoovqaalvaaNo ratings yet
- Bioquimica Sistema DigestivoDocument26 pagesBioquimica Sistema DigestivojesusmonNo ratings yet
- Hipertensión Arterial SistemicaDocument32 pagesHipertensión Arterial SistemicaMiguel Avalos100% (3)
- Resumen 4 Control Del DolorDocument20 pagesResumen 4 Control Del DolorMireya OsunaNo ratings yet
- Atp y Vias de Señalizacion CelularDocument4 pagesAtp y Vias de Señalizacion CelularMarq TintaNo ratings yet
- t8 Situaciones EspecialesDocument7 pagest8 Situaciones EspecialesLucia Fernanda Rivera ChavezNo ratings yet
- Tomo Vi - LetrasDocument132 pagesTomo Vi - LetrasJhon MezaNo ratings yet
- Ficha de Monitoreo PROGRAMACION 2015Document10 pagesFicha de Monitoreo PROGRAMACION 2015Anonymous N9EBVnbNo ratings yet
- Salud y EnfermedadDocument13 pagesSalud y EnfermedadAnonymous b7BELE0% (1)
- Proyecto Sobre El MalojilloDocument12 pagesProyecto Sobre El Malojilloysbecarmen60% (5)