You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5784)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (72)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Risks of cancer and mental health issues from drinkingDocument3 pagesRisks of cancer and mental health issues from drinkingRochel LescanoNo ratings yet
- Electrical InjuriesDocument1 pageElectrical InjuriesIkhwan SalamNo ratings yet
- Metformin A Review of Its Use in The Treatment Typ-DikonversiDocument30 pagesMetformin A Review of Its Use in The Treatment Typ-DikonversiAida H.djamhuriNo ratings yet
- Ventouse DeliveryDocument43 pagesVentouse DeliveryN. Siva100% (1)
- Abdominal Trauma PDFDocument17 pagesAbdominal Trauma PDFCarlos UrquijoNo ratings yet
- Package Leaflet: Information For The User Cetirizine Hydrochloride 5 MG/ 5 ML Oral SolutionDocument7 pagesPackage Leaflet: Information For The User Cetirizine Hydrochloride 5 MG/ 5 ML Oral SolutionZulva Chairunnisa BudimanNo ratings yet
- CBC reference ranges and abnormal conditionsDocument4 pagesCBC reference ranges and abnormal conditionsndrfernandezNo ratings yet
- Department of HealthDocument2 pagesDepartment of HealthdenNo ratings yet
- Garp-Nepal - Sa (Global Antibiotic Resistance Partnership)Document90 pagesGarp-Nepal - Sa (Global Antibiotic Resistance Partnership)Nivea VazNo ratings yet
- Caffeine Intoxication Consumer MonographDocument3 pagesCaffeine Intoxication Consumer MonographEfdio Manuel Joshiro SiagianNo ratings yet
- Bad & BSF Sunscreen Fact SheetDocument9 pagesBad & BSF Sunscreen Fact SheetArkadiyNo ratings yet
- Emergency TriageDocument8 pagesEmergency TriageJam Chelsea ChyNo ratings yet
- Apoptosis - An Introduction For The Endodontist PDFDocument9 pagesApoptosis - An Introduction For The Endodontist PDFRamona MateiNo ratings yet
- April 2017-1 MJDFDocument24 pagesApril 2017-1 MJDFsajna1980No ratings yet
- Approach To Child With Fever: Liew Qian YiDocument33 pagesApproach To Child With Fever: Liew Qian YinavenNo ratings yet
- Neuropathology of Autonomic Dysfunction in SynucleinopathiesDocument10 pagesNeuropathology of Autonomic Dysfunction in SynucleinopathiesAnna KolpinaNo ratings yet
- Kewal K. Jain - Textbook of Personalized Medicine-Springer (2021)Document770 pagesKewal K. Jain - Textbook of Personalized Medicine-Springer (2021)youssraNo ratings yet
- Oxyganation: Is IsDocument5 pagesOxyganation: Is IsRahmatun NurainaNo ratings yet
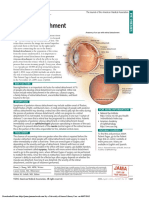
- Retinal Detachment: Jama Patient PageDocument1 pageRetinal Detachment: Jama Patient PageKemal TaufikNo ratings yet
- ĐỀ 1 MH2022.HSDocument6 pagesĐỀ 1 MH2022.HShong hieu NguyenNo ratings yet
- Codex OccultusDocument4 pagesCodex OccultusCaroline Erstwhile100% (1)
- Introduction of BMEDocument7 pagesIntroduction of BMEmuhammadfawadnNo ratings yet
- 2019, Volume 6, No 1: ISSN 2397-5628Document46 pages2019, Volume 6, No 1: ISSN 2397-5628djonoumaNo ratings yet
- History Taking and Physical Examination in Obstetrics and GynecologyDocument42 pagesHistory Taking and Physical Examination in Obstetrics and GynecologyAhlam Azam, MohamedNo ratings yet
- Indriyaaneekam of Charaka Indriya Sthana - An Explorative StudyDocument13 pagesIndriyaaneekam of Charaka Indriya Sthana - An Explorative StudyPrasad MamidiNo ratings yet
- Cardiovascular Disease in Rheumatoid Arthritis: Single-Center Hospital-Based Cohort Study in FranceDocument15 pagesCardiovascular Disease in Rheumatoid Arthritis: Single-Center Hospital-Based Cohort Study in Francemarisa isahNo ratings yet
- THE Quality OF Life AND Caregiving Burden Among Caregivers OF People With Dementia IN Hanoi, BAC Ninh AND HAI Phong, VietnamDocument310 pagesTHE Quality OF Life AND Caregiving Burden Among Caregivers OF People With Dementia IN Hanoi, BAC Ninh AND HAI Phong, VietnamCHEE KOK SENGNo ratings yet
- Top 15 Anti-Inflammatory FoodsDocument7 pagesTop 15 Anti-Inflammatory FoodsAli HassanNo ratings yet
- Symptoms Board Game EslDocument4 pagesSymptoms Board Game EslNguyễn LinhNo ratings yet
- Acute Respiratory Distress Syndrome (ARDS)Document85 pagesAcute Respiratory Distress Syndrome (ARDS)Jose Adan Vazquez MoralesNo ratings yet