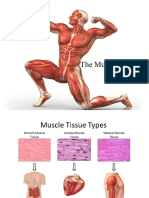
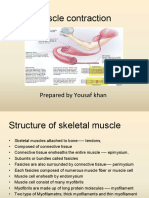
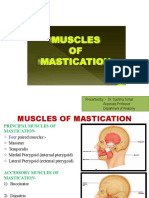
You might also like
- Skeletal MuscleDocument98 pagesSkeletal MuscleMoon Moon100% (1)
- Muscle PhysiologyDocument24 pagesMuscle PhysiologyArmando AlehandroNo ratings yet
- How Muscles Generate MovementDocument38 pagesHow Muscles Generate Movementtoyy100% (1)
- Muscle DentalDocument74 pagesMuscle Dentalapi-3710331No ratings yet
- Muscle Pgy - 222Document48 pagesMuscle Pgy - 222SAVIOUR BANDANo ratings yet
- Contractile Cells: Skeletal Muscle Structure and FunctionDocument116 pagesContractile Cells: Skeletal Muscle Structure and Functionnav_malhiNo ratings yet
- 6.muscular TissueDocument40 pages6.muscular Tissuemero wowNo ratings yet
- 3D. Muscle TissueDocument47 pages3D. Muscle Tissuejaf123xyNo ratings yet
- Nerve and Muscle Lecture 6 paraDocument24 pagesNerve and Muscle Lecture 6 paraTatenda SibandaNo ratings yet
- SM Skeletal Muscle SlidesDocument68 pagesSM Skeletal Muscle Slideskarishma.7022No ratings yet
- Muscle Intro-Skeletal Muscle IDocument31 pagesMuscle Intro-Skeletal Muscle IPehovelo JohannesNo ratings yet
- Muscular System: Histology and PhysiologyDocument36 pagesMuscular System: Histology and PhysiologyFajar YuliantoNo ratings yet
- Skeletal MusclesDocument7 pagesSkeletal MusclesAhmed OudahNo ratings yet
- Microscopic Anatomy of A Skeletal Muscle FiberDocument33 pagesMicroscopic Anatomy of A Skeletal Muscle FiberKumar MSNo ratings yet
- Physiology Slide #1Document18 pagesPhysiology Slide #1anon_974423120No ratings yet
- Muscles and Muscle Tissue: Muscle Overview Muscle SimilaritiesDocument11 pagesMuscles and Muscle Tissue: Muscle Overview Muscle SimilaritiesHerne BalberdeNo ratings yet
- Chapter 10: Muscle Tissue: VoluntaryDocument13 pagesChapter 10: Muscle Tissue: VoluntaryClyde BaltazarNo ratings yet
- Histology of Cardiac and Smooth MuscleDocument73 pagesHistology of Cardiac and Smooth MuscleShehryar AbbasNo ratings yet
- Histology of Cardiac and Smooth MuscleDocument73 pagesHistology of Cardiac and Smooth MuscleShehryar AbbasNo ratings yet
- Excitation Contraction Coupling: Nandini GoyalDocument32 pagesExcitation Contraction Coupling: Nandini GoyalNandini GoyalNo ratings yet
- Contraction of Skeletal Muscle (1) : Narrated LectureDocument45 pagesContraction of Skeletal Muscle (1) : Narrated LectureDeanNo ratings yet
- Muscle Tissue: - Skeletal Muscle - Cardiac Muscle - Smooth MuscleDocument36 pagesMuscle Tissue: - Skeletal Muscle - Cardiac Muscle - Smooth MuscleDentist Dina Samy100% (1)
- Histology 6 MuscleDocument88 pagesHistology 6 MuscleAbdul RahmanNo ratings yet
- Health PPT 3 - Muscle 2013-1Document50 pagesHealth PPT 3 - Muscle 2013-1Mengistu GebeyehuNo ratings yet
- MUSCLE CELL PHYSIOLOGY MBCHB YEAR 1 2016-2Document289 pagesMUSCLE CELL PHYSIOLOGY MBCHB YEAR 1 2016-2AdminNo ratings yet
- MUSCLE PHYSIOLOGY: AN OVERVIEWDocument50 pagesMUSCLE PHYSIOLOGY: AN OVERVIEWJoyce Adjei-boateng100% (1)
- Unit 3 Muscle Physiology - BPT PDFDocument55 pagesUnit 3 Muscle Physiology - BPT PDFRadhikaNo ratings yet
- 4.1 PhysDocument27 pages4.1 PhysshivaniNo ratings yet
- Sarcolemma - Cell MembraneDocument66 pagesSarcolemma - Cell MembraneAman SyedNo ratings yet
- Muscle Lecture On (Excitable Tissue)Document126 pagesMuscle Lecture On (Excitable Tissue)Muhammed ZainabNo ratings yet
- Muscle Tissue-Wl06.11Document54 pagesMuscle Tissue-Wl06.11api-19641337No ratings yet
- The Muscular System: Types, Structure & FunctionDocument50 pagesThe Muscular System: Types, Structure & FunctionArina GainaNo ratings yet
- Physiology of Skeletal & Smooth MusclesDocument10 pagesPhysiology of Skeletal & Smooth MusclesAli AlhamadNo ratings yet
- Chapter 7 - Muscular SystemDocument29 pagesChapter 7 - Muscular SystemlNo ratings yet
- Muscle Tissue NotesDocument11 pagesMuscle Tissue NotesAubrey Unique EvangelistaNo ratings yet
- Muscle Tissue LectureDocument34 pagesMuscle Tissue LectureurjaNo ratings yet
- Anaphy-Muscular HandoutDocument7 pagesAnaphy-Muscular HandoutAEGEL DARO TORRALBANo ratings yet
- Lec4 Muscles CompressedDocument27 pagesLec4 Muscles Compressedmohammedbaker1q2w3eNo ratings yet
- Actin Myosin CytoskeletonDocument28 pagesActin Myosin Cytoskeletonjesus rajeshdivyaNo ratings yet
- General Mechanism of Muscle ContractionDocument22 pagesGeneral Mechanism of Muscle ContractionAnuya Kulkarni-DeoNo ratings yet
- Muscle Physiology ExplainedDocument24 pagesMuscle Physiology Explainedroqaia roqaiaNo ratings yet
- Skeletal Muscle Physiology ExplainedDocument26 pagesSkeletal Muscle Physiology ExplainedUSAMA AHMEDNo ratings yet
- Muscular System Explained: Skeletal, Cardiac & Smooth Muscle TissueDocument44 pagesMuscular System Explained: Skeletal, Cardiac & Smooth Muscle TissueClaudine Jo B. TalabocNo ratings yet
- Lecture 1 Physiology of Skeletal Muscle by Dr. RoomiDocument37 pagesLecture 1 Physiology of Skeletal Muscle by Dr. RoomiMudassar Roomi100% (1)
- MuscleDocument11 pagesMuscleJeff ParkNo ratings yet
- Muscles MidtermDocument32 pagesMuscles MidtermMata, Ersel MaeNo ratings yet
- Muscular SystemDocument10 pagesMuscular SystemMarie Veatrice JacomilleNo ratings yet
- Azantee Yazmie Abdul Wahab Dept. of Obstetrics & Gynaecology Kulliyyah of Medicine IIUM KuantanDocument51 pagesAzantee Yazmie Abdul Wahab Dept. of Obstetrics & Gynaecology Kulliyyah of Medicine IIUM KuantanNur ShafilaNo ratings yet
- NotesDocument15 pagesNotesGizem OsmanogluNo ratings yet
- Contraction of Skeletal Muscle.Document19 pagesContraction of Skeletal Muscle.Salman KhanNo ratings yet
- Kharkov National Medical University: Muscle TissueDocument42 pagesKharkov National Medical University: Muscle TissueSyed Irfan ArifNo ratings yet
- Skeletal Muscle Classification and StructureDocument43 pagesSkeletal Muscle Classification and StructureHusnain WattoNo ratings yet
- The Muscular System: DR Faisal Institute of Physiology and PharmacologyDocument61 pagesThe Muscular System: DR Faisal Institute of Physiology and PharmacologyTask BirdNo ratings yet
- Muscle Physiology in Orthodontics / Orthodontic Courses by Indian Dental AcademyDocument50 pagesMuscle Physiology in Orthodontics / Orthodontic Courses by Indian Dental Academyindian dental academyNo ratings yet
- Anatomy and Physiology Muscular System OverviewDocument7 pagesAnatomy and Physiology Muscular System OverviewKei GauranoNo ratings yet
- MuscularDocument61 pagesMuscularFranz DiazNo ratings yet
- Muscle: Gerald Dale Giron MD Department of Human Structural BiologyDocument23 pagesMuscle: Gerald Dale Giron MD Department of Human Structural BiologyMarx AsuncionNo ratings yet
- Wa0038Document10 pagesWa0038Shehzad QureshiNo ratings yet
- The Basics of Cell Life with Max Axiom, Super Scientist: 4D An Augmented Reading Science ExperienceFrom EverandThe Basics of Cell Life with Max Axiom, Super Scientist: 4D An Augmented Reading Science ExperienceNo ratings yet
- Gametogenesis 1Document10 pagesGametogenesis 1Affie SaikolNo ratings yet
- Gametogenesis - Trans PDFDocument9 pagesGametogenesis - Trans PDFAffie SaikolNo ratings yet
- The Epithelial CellsDocument5 pagesThe Epithelial CellsAffie SaikolNo ratings yet
- Microbio Questions CompilationDocument21 pagesMicrobio Questions CompilationAffie SaikolNo ratings yet
- Epithelial TissueDocument62 pagesEpithelial TissueAffie SaikolNo ratings yet
- Microbiology - NotesDocument1 pageMicrobiology - NotesAffie SaikolNo ratings yet
- 02 BiomoleculesDocument17 pages02 BiomoleculesAffie SaikolNo ratings yet
- Histology of the larynx, bronchi, and lungsDocument49 pagesHistology of the larynx, bronchi, and lungsAffie SaikolNo ratings yet
- Connective TissueDocument74 pagesConnective TissueAffie SaikolNo ratings yet
- 01 Intro To Biochem - EditDocument5 pages01 Intro To Biochem - EditJoanne AjosNo ratings yet
- COPD Patient With Chronic Dyspnea and CyanosisDocument2 pagesCOPD Patient With Chronic Dyspnea and CyanosisAffie SaikolNo ratings yet
- Gametogenesis - Trans PDFDocument9 pagesGametogenesis - Trans PDFAffie SaikolNo ratings yet
- Delfin, RMTDocument34 pagesDelfin, RMTAffie SaikolNo ratings yet
- Recall 1Document4 pagesRecall 1pikachuNo ratings yet
- Antigen Antibody Reaction (MUST READ)Document137 pagesAntigen Antibody Reaction (MUST READ)Affie SaikolNo ratings yet
- Autoimmune DiseasesDocument35 pagesAutoimmune DiseasesAffie SaikolNo ratings yet
- Lesson Korean NumbersDocument5 pagesLesson Korean NumbersLulu Afifah OctaviaNo ratings yet
- Serological Diagnosis of Bacterial InfectionsDocument81 pagesSerological Diagnosis of Bacterial InfectionsAffie SaikolNo ratings yet
- Hindi Phase1 BKLTDocument32 pagesHindi Phase1 BKLTSam KuttyNo ratings yet
- Prayer Times in Vigan (Ilocos) - Year 2018: PrintDocument2 pagesPrayer Times in Vigan (Ilocos) - Year 2018: PrintAffie SaikolNo ratings yet
- HomeostasisDocument3 pagesHomeostasisAffie SaikolNo ratings yet
- Denr OfficialDocument1 pageDenr OfficialAffie SaikolNo ratings yet
- Understanding IslamDocument31 pagesUnderstanding IslamAffie SaikolNo ratings yet
- Introduction Ana and PhysioDocument54 pagesIntroduction Ana and PhysioAffie SaikolNo ratings yet
- Environmental Ethics in IslamDocument74 pagesEnvironmental Ethics in IslamAffie SaikolNo ratings yet
- Electron Transport ChainDocument27 pagesElectron Transport ChainAffie SaikolNo ratings yet
- AnatomyDocument52 pagesAnatomyAffie SaikolNo ratings yet
- Cardiovascular SystemDocument64 pagesCardiovascular SystemAffie SaikolNo ratings yet
- Siklus Asam Sitrat PDFDocument27 pagesSiklus Asam Sitrat PDFLuses Shantia HaryantoNo ratings yet
- Turbidimetry and Nephelometry1Document3 pagesTurbidimetry and Nephelometry1maxim_crank6101100% (1)
- Nervous Responses of EarthwormDocument27 pagesNervous Responses of EarthwormKayla JoezetteNo ratings yet
- LWT - Food Science and TechnologyDocument8 pagesLWT - Food Science and TechnologyAzb 711No ratings yet
- Flowering PlantsDocument43 pagesFlowering Plantskingbanakon100% (1)
- Cooling Tower Operation - 030129Document8 pagesCooling Tower Operation - 030129Sekar CmNo ratings yet
- Reproductive Biology and Phylogeny of Birds PDFDocument624 pagesReproductive Biology and Phylogeny of Birds PDFFabricio GZNo ratings yet
- Lesson 7 - Organ TransplantationDocument14 pagesLesson 7 - Organ TransplantationJosee100% (1)
- Introduction To Bioinformatics: Database Search (FASTA)Document35 pagesIntroduction To Bioinformatics: Database Search (FASTA)mahedi hasanNo ratings yet
- Eagles and Eaglets gr2Document2 pagesEagles and Eaglets gr2vovanmtcNo ratings yet
- LearnerDocument7 pagesLearnersudhacarhrNo ratings yet
- 03 Chapter 3Document148 pages03 Chapter 3arghaNo ratings yet
- WCH03 01 Que 20180124Document16 pagesWCH03 01 Que 20180124Rameez Mazhar SiddiqiNo ratings yet
- MDU Open Elective 3rd Sem Date Sheet May 2019Document1 pageMDU Open Elective 3rd Sem Date Sheet May 2019Ratan DeshNo ratings yet
- Correlation of Dynamic Strength in The Standing Calf Raise With Sprinting Performance in Consecutive Sections Up To 30 MetersDocument9 pagesCorrelation of Dynamic Strength in The Standing Calf Raise With Sprinting Performance in Consecutive Sections Up To 30 MetersJohn NixonNo ratings yet
- Perkecambahan Dan Pertumbuhan Palem Jepang (Actinophloeus Macarthurii Becc.) Akibat Perendaman Biji Dalam LumpurDocument7 pagesPerkecambahan Dan Pertumbuhan Palem Jepang (Actinophloeus Macarthurii Becc.) Akibat Perendaman Biji Dalam LumpurNurma YahyaNo ratings yet
- Physiology of Cerebellum Lec Foe 2nd Year by DR Sadia Uploaded by ZaighamDocument61 pagesPhysiology of Cerebellum Lec Foe 2nd Year by DR Sadia Uploaded by ZaighamZaigham HammadNo ratings yet
- Zoologija VrsteDocument16 pagesZoologija VrsteДушан МарковићNo ratings yet
- Science10 Quarter4 Week3-BiomoleculesDocument2 pagesScience10 Quarter4 Week3-BiomoleculesMargareth LandichoNo ratings yet
- 4 Factors Effecting Shelf Life and Storage Life of Fresh FishDocument26 pages4 Factors Effecting Shelf Life and Storage Life of Fresh FishRafidah Zakaria100% (1)
- Botany AssignmentDocument35 pagesBotany AssignmentLakshmiNo ratings yet
- Protochordata FIXDocument33 pagesProtochordata FIXSylvia AnggraeniNo ratings yet
- Metals: Methods To Evaluate Corrosion in Buried Steel Structures: A ReviewDocument21 pagesMetals: Methods To Evaluate Corrosion in Buried Steel Structures: A ReviewPeymanMajidiNo ratings yet
- Muscle Energy Techniques ExplainedDocument2 pagesMuscle Energy Techniques ExplainedAtanu Datta100% (1)
- Presented By:-Dr. Sushma Tomar Associate Professor Department of AnatomyDocument20 pagesPresented By:-Dr. Sushma Tomar Associate Professor Department of AnatomyCristine EchaveNo ratings yet
- Kenya's Natural Capital AtlasDocument139 pagesKenya's Natural Capital AtlasElvis Ole Sankale Ntimama100% (7)
- The Anatomy, Physiology, and Pathophysiology of Muscular SystemDocument55 pagesThe Anatomy, Physiology, and Pathophysiology of Muscular SystemAnisaNo ratings yet
- Ewh Ix PDFDocument80 pagesEwh Ix PDFOR Premium FreeNo ratings yet
- Cell Division GizmoDocument3 pagesCell Division Gizmoapi-522847737No ratings yet
- Trunking CatalogDocument8 pagesTrunking CatalogDenuka PathiranaNo ratings yet
- Cu EnzymesDocument8 pagesCu EnzymesVINEESHA VKNo ratings yet