You might also like
- Manejo fitosanitario del cultivo de hortalizas medidas para la temporada invernalFrom EverandManejo fitosanitario del cultivo de hortalizas medidas para la temporada invernalNo ratings yet
- Algunas especies de moscas de la fruta, exóticas para ColombiaFrom EverandAlgunas especies de moscas de la fruta, exóticas para ColombiaNo ratings yet
- DiapositivasDocument10 pagesDiapositivasFernan FloresNo ratings yet
- Bt insecticida naturalDocument20 pagesBt insecticida naturalMarllybi MarinNo ratings yet
- IsabionDocument3 pagesIsabionciaraNo ratings yet
- La Palomilla Dorso de DiamanteDocument7 pagesLa Palomilla Dorso de DiamanteaaronNo ratings yet
- Metabolitos Secundarios en Trichoderma Spp. y Sus AplicacionesDocument3 pagesMetabolitos Secundarios en Trichoderma Spp. y Sus Aplicacionesyamilet yucraNo ratings yet
- Toma Muestras Diagnóstico FitosanitarioDocument20 pagesToma Muestras Diagnóstico FitosanitarioCALF86% (7)
- Control BiologicoDocument228 pagesControl BiologicoJORGE ROQUE AREVALO100% (1)
- Folleto Maiz 11Document20 pagesFolleto Maiz 11aprendiz777No ratings yet
- Poster Trichoderma Control PestalotiaDocument1 pagePoster Trichoderma Control PestalotiaJessica Rodriguez EscobarNo ratings yet
- Informe Identificacion Plagas de CitricosDocument15 pagesInforme Identificacion Plagas de CitricosVERYK MOREYNo ratings yet
- Control-BiologicooDocument26 pagesControl-BiologicooJuanCarminPeñaNo ratings yet
- Cultivo de HortalizasDocument51 pagesCultivo de HortalizaslindaaquinoNo ratings yet
- Manejo Integrado de Plagas IiDocument22 pagesManejo Integrado de Plagas IiYanderin Cecilia Marcos ChampionNo ratings yet
- Módulo 4. Alternativas Al Uso de PesticidasDocument55 pagesMódulo 4. Alternativas Al Uso de PesticidasJACB 69No ratings yet
- Bacterias FitopatógenasDocument46 pagesBacterias Fitopatógenasedwin100% (1)
- Manejo Agroecologico Del Gusano Cogollero Del Maiz en MexicoDocument10 pagesManejo Agroecologico Del Gusano Cogollero Del Maiz en MexicoasmanctNo ratings yet
- Efectos Nocivos Provocados Por El Uso de PlaguicidasDocument21 pagesEfectos Nocivos Provocados Por El Uso de PlaguicidasarchieNo ratings yet
- Aplicación Del Control Biológico en El Cultivo de La Palta Como Parte Del Manejo Integrado de PlagasDocument4 pagesAplicación Del Control Biológico en El Cultivo de La Palta Como Parte Del Manejo Integrado de Plagasdalmert50% (2)
- Control Genetico de Plagas AgricolasDocument17 pagesControl Genetico de Plagas AgricolasHECTOR VELASQUEZ ALCANTARANo ratings yet
- Riego en FrutalesDocument58 pagesRiego en FrutalesCarolina Soledad Ordenes VàsquezNo ratings yet
- Virus FitopatógenosDocument29 pagesVirus FitopatógenosGiannina Campoverde VenturaNo ratings yet
- Bacilus Subtilis Sobre ClavibacterDocument36 pagesBacilus Subtilis Sobre ClavibacterRudy Altamirano RuizNo ratings yet
- Cria de Insectos Plaga para El MIP Quinua - R QuispeDocument8 pagesCria de Insectos Plaga para El MIP Quinua - R QuispePamelaSantosNo ratings yet
- Control de Plagas y EnfermedadesDocument6 pagesControl de Plagas y EnfermedadesMarioZapetaNo ratings yet
- Producción, Uso y Control de Calidad de HONGOS ENTOMOPATOGENOSDocument9 pagesProducción, Uso y Control de Calidad de HONGOS ENTOMOPATOGENOSAsura NephilimNo ratings yet
- Inoculos MicrobianosDocument19 pagesInoculos MicrobianosEnrique Ruiz TapiaNo ratings yet
- Hongos Entomopatogenos de Bemisia TabaciDocument18 pagesHongos Entomopatogenos de Bemisia TabaciWilberth Chan Cupul100% (1)
- 1 B Cuestionario Del MIP-1Document2 pages1 B Cuestionario Del MIP-1Alexander AnguloNo ratings yet
- Insectos plaga agrícolaDocument144 pagesInsectos plaga agrícolaThaily SanchezNo ratings yet
- Variedades y Mejoramiento Genetico Del CafeDocument5 pagesVariedades y Mejoramiento Genetico Del CafeGamus del PeruNo ratings yet
- 1 Mohos 2015 PDFDocument41 pages1 Mohos 2015 PDFThewilson H. RamosNo ratings yet
- Control insectos alfalfaDocument25 pagesControl insectos alfalfaGladys Luna HerreraNo ratings yet
- Plagas y enfermedades del maízDocument17 pagesPlagas y enfermedades del maízJorge LuisNo ratings yet
- Fitopatologia Agricola 2020-IiDocument11 pagesFitopatologia Agricola 2020-IiAlex CuriNo ratings yet
- Hongos Entomófagos - Docx TrabajoDocument14 pagesHongos Entomófagos - Docx TrabajoLeonel Silvestre CardonaNo ratings yet
- Entomologia, Microbiologia, Fitopatologia y MalherbologiaDocument35 pagesEntomologia, Microbiologia, Fitopatologia y MalherbologiaEliceo Ortiz Vergara67% (3)
- Practica de Entomologia Agricola Rinconazo 2017Document9 pagesPractica de Entomologia Agricola Rinconazo 2017EDSON MONTENEGRO RODRIGUEZNo ratings yet
- Clases FitopatologíaDocument39 pagesClases FitopatologíaGabriel Tapia100% (1)
- Manejo Integrado de Plagas Insectiles y MipDocument50 pagesManejo Integrado de Plagas Insectiles y MipIsmaelNo ratings yet
- Informe Virosis en PapaDocument2 pagesInforme Virosis en PapaJ Luis Inga ArmijosNo ratings yet
- Cuestionario Programa Control de Plagas PrintoglassDocument23 pagesCuestionario Programa Control de Plagas PrintoglassAlexxAnderCrIstianaNo ratings yet
- Manualde Producciondel Cultivode Cacahua PDFDocument86 pagesManualde Producciondel Cultivode Cacahua PDFJose Melchor MarroquinNo ratings yet
- La Fenologia Del Maiz y Su Relacion Con La Incidencia de PlagasDocument3 pagesLa Fenologia Del Maiz y Su Relacion Con La Incidencia de PlagasFaustino Ramirez75% (4)
- Convertido en Word Hidroponia de La UnschDocument96 pagesConvertido en Word Hidroponia de La UnschDeisy Mitma Enciso100% (1)
- Producción y Uso de Trichogramma en Anzoátegui, VenezuelaDocument3 pagesProducción y Uso de Trichogramma en Anzoátegui, VenezuelaRamón GarcíaNo ratings yet
- ControlLepidópterosBacillus40Document2 pagesControlLepidópterosBacillus40navaron64No ratings yet
- Control de Malezas 2Document59 pagesControl de Malezas 2Claudia Vera BenitesNo ratings yet
- Semana 5 Practicas AgronomicasDocument21 pagesSemana 5 Practicas AgronomicasYezy Huaman MosqueraNo ratings yet
- Hongos Control PlagasDocument73 pagesHongos Control PlagasmorenoandyNo ratings yet
- Manejo Integrado de Tranzschelia DiscolorDocument18 pagesManejo Integrado de Tranzschelia DiscolorJonathan Chugnas Chuquilin100% (1)
- Nematodos FitoparasitosDocument13 pagesNematodos FitoparasitosJean Paul Alvarez MartinezNo ratings yet
- Mildiu de La VidDocument11 pagesMildiu de La VidTalia Janeth Soto TéllezNo ratings yet
- Mancha Foliares ApazaDocument28 pagesMancha Foliares ApazaCleynthon Chanel Huayra CrispinNo ratings yet
- Plagas CitricosDocument29 pagesPlagas CitricosMario Argandoña DominguezNo ratings yet
- ENTOMOPATÓGENOSDocument9 pagesENTOMOPATÓGENOSGrover Camasca AronesNo ratings yet
- Plantas Indicadoras Quincho Vera Semana 10Document8 pagesPlantas Indicadoras Quincho Vera Semana 10Sandro CruzNo ratings yet
- Examen de Entra Estructuras HidraulicasDocument3 pagesExamen de Entra Estructuras HidraulicasBarreto Noa Dany DarioNo ratings yet
- Examen de HidraulicaDocument4 pagesExamen de HidraulicaBarreto Noa Dany DarioNo ratings yet
- La Ética Ambiental Como Reflexión: Hacia El Desarrollo de La Conciencia de La ResponsabilidadDocument7 pagesLa Ética Ambiental Como Reflexión: Hacia El Desarrollo de La Conciencia de La ResponsabilidadMiguel CMNo ratings yet
- Practica de Rapidas BARRETO NOADocument4 pagesPractica de Rapidas BARRETO NOABarreto Noa Dany DarioNo ratings yet
- EXAMEN DE HIDRAULICA Barreto Noa Dario PDFDocument4 pagesEXAMEN DE HIDRAULICA Barreto Noa Dario PDFBarreto Noa Dany DarioNo ratings yet
- Practica de Rapidas BARRETO NOADocument4 pagesPractica de Rapidas BARRETO NOABarreto Noa Dany DarioNo ratings yet
- Examen 1 Ejercicio 2Document1 pageExamen 1 Ejercicio 2Barreto Noa Dany DarioNo ratings yet
- Tema03 - Ley de DarcyDocument24 pagesTema03 - Ley de DarcyCarol Yuna AlcaNo ratings yet
- Resumen 33-41Document8 pagesResumen 33-41Barreto Noa Dany DarioNo ratings yet
- Diseño de SedimentadoresDocument20 pagesDiseño de SedimentadoresBarreto Noa Dany DarioNo ratings yet
- Ética, moral y educación en la formación de valoresDocument3 pagesÉtica, moral y educación en la formación de valoresBarreto Noa Dany DarioNo ratings yet
- Barreto Noa Dario PDFDocument6 pagesBarreto Noa Dario PDFBarreto Noa Dany DarioNo ratings yet
- Examen Ejercicio 1Document2 pagesExamen Ejercicio 1Barreto Noa Dany DarioNo ratings yet
- Etica CorrupcionDocument3 pagesEtica CorrupcionBarreto Noa Dany DarioNo ratings yet
- Copia de Practica1 - FormatoDocument8 pagesCopia de Practica1 - FormatoLeonar Bojorges CoariteNo ratings yet
- Cómo Calcular El Flujo de Caja LibreDocument2 pagesCómo Calcular El Flujo de Caja LibreBarreto Noa Dany DarioNo ratings yet
- Universidad Nacional de San Agustín de ArequipaDocument20 pagesUniversidad Nacional de San Agustín de ArequipaBarreto Noa Dany DarioNo ratings yet
- Universidad Nacional de San Agustín de ArequipaDocument20 pagesUniversidad Nacional de San Agustín de ArequipaBarreto Noa Dany DarioNo ratings yet
- Examen de HidraulicaDocument4 pagesExamen de HidraulicaBarreto Noa Dany DarioNo ratings yet
- Barreto No Dario Examen 2 PDFDocument5 pagesBarreto No Dario Examen 2 PDFBarreto Noa Dany DarioNo ratings yet
- Barreto Noa Dario PDFDocument6 pagesBarreto Noa Dario PDFBarreto Noa Dany DarioNo ratings yet
- Barreto No Dario Examen 2 PDFDocument5 pagesBarreto No Dario Examen 2 PDFBarreto Noa Dany DarioNo ratings yet
- EXAMEN DE HIDRAULICA Barreto Noa Dario PDFDocument4 pagesEXAMEN DE HIDRAULICA Barreto Noa Dario PDFBarreto Noa Dany DarioNo ratings yet
- 01e Und Cap1 1703252-Imic Epis 2020BDocument5 pages01e Und Cap1 1703252-Imic Epis 2020BBarreto Noa Dany DarioNo ratings yet
- Ex EntradaDocument2 pagesEx EntradaBarreto Noa Dany DarioNo ratings yet
- Ley y Reglam. ActualDocument88 pagesLey y Reglam. ActualLeslieCoronelNo ratings yet
- ResultadosDocument1 pageResultadosBarreto Noa Dany DarioNo ratings yet
- Seminario 1 Tratamiendo de Aguas BARRETO NOADocument13 pagesSeminario 1 Tratamiendo de Aguas BARRETO NOABarreto Noa Dany DarioNo ratings yet
- N° 7Document9 pagesN° 7Barreto Noa Dany DarioNo ratings yet
- Ley y Reglam. ActualDocument88 pagesLey y Reglam. ActualLeslieCoronelNo ratings yet
- 1.08 EXAMEN FÍSICO DEL RECIÉN NACIDO RespuestasDocument4 pages1.08 EXAMEN FÍSICO DEL RECIÉN NACIDO RespuestasJavier VelasteguiNo ratings yet
- Triptico Proteccion AuditivaDocument4 pagesTriptico Proteccion AuditivaEdil AguilarNo ratings yet
- Plan de Almacenamiento Sustancias QuimicasDocument18 pagesPlan de Almacenamiento Sustancias QuimicasNelson CifuentesNo ratings yet
- Lineamientos Técnicos para La Prevención y Control de RabiaDocument30 pagesLineamientos Técnicos para La Prevención y Control de RabiaJorge Raul Avalos ValenciaNo ratings yet
- Ácido glutámico: el aminoácido clave para la salud cerebral y muscularDocument3 pagesÁcido glutámico: el aminoácido clave para la salud cerebral y muscularKevin Cáceres ContrerasNo ratings yet
- Onfalitis y Conjuntivitis NeonatalDocument17 pagesOnfalitis y Conjuntivitis Neonatalcinthia cardenasNo ratings yet
- Amor eterno de Bolívar y ManuelitaDocument3 pagesAmor eterno de Bolívar y Manuelitafparra27No ratings yet
- Prospecto Web Dipifebral 03Document1 pageProspecto Web Dipifebral 03Débora IbarrolaNo ratings yet
- Diapositivas de La Propuesta de Un Sistema de Seguridad y Salud para Una Empresa AgroindustrialDocument64 pagesDiapositivas de La Propuesta de Un Sistema de Seguridad y Salud para Una Empresa AgroindustrialEnrique GuyinNo ratings yet
- Antecedentes Del Tratamiento Con AnimalesDocument37 pagesAntecedentes Del Tratamiento Con Animalesruben.solisescNo ratings yet
- Semiología de Los Sindromes MotoresDocument20 pagesSemiología de Los Sindromes Motoresnaranja tazaNo ratings yet
- Guia Basica DR Benjamin PHDDocument12 pagesGuia Basica DR Benjamin PHDMariella HdezNo ratings yet
- Cuadernillo N°4 (Ciencias 7°) ReproduccionDocument31 pagesCuadernillo N°4 (Ciencias 7°) ReproduccionFrancisco Osvaldo Hernández LópezNo ratings yet
- Microbiologia EstomatológicaDocument24 pagesMicrobiologia EstomatológicaPAúl Casa OrtizNo ratings yet
- Hipertension ArterialDocument15 pagesHipertension ArterialSergio GonzalesNo ratings yet
- Guía Med Trabajo 2do Parcial PDFDocument32 pagesGuía Med Trabajo 2do Parcial PDFGuillermo EsparzzaNo ratings yet
- Hematuria y ProteinuriaDocument12 pagesHematuria y ProteinuriaAlex RojasNo ratings yet
- Meningitis tuberculosa: una emergencia médicaDocument10 pagesMeningitis tuberculosa: una emergencia médicaRAUL ROSALES HNo ratings yet
- Prevenimss PresentacionDocument23 pagesPrevenimss PresentacionMague Fabela Lozano100% (1)
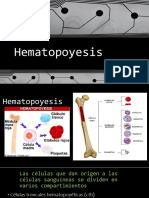
- Hematopoyesis FisiologiaDocument18 pagesHematopoyesis FisiologiaJuan Felipe PadillaNo ratings yet
- Taller 12 Armas Quimicas y NuclearesDocument6 pagesTaller 12 Armas Quimicas y Nucleareslorieth barriosNo ratings yet
- Hipertensión IntracraneanaDocument2 pagesHipertensión IntracraneanaDavid PuyóNo ratings yet
- Pruebas de Lab en Gestantes - Mblgo Jose CH PDFDocument21 pagesPruebas de Lab en Gestantes - Mblgo Jose CH PDFPaul VeasNo ratings yet
- Asma AdultosDocument13 pagesAsma AdultosalejandroNo ratings yet
- LISTA DE CHEQUEO RESOLUCION 0312 DE 2019 TornoDocument53 pagesLISTA DE CHEQUEO RESOLUCION 0312 DE 2019 TornoBeatriz Elena SALINAS ORDONEZ87% (15)
- Defectos de Los OvinosDocument31 pagesDefectos de Los OvinosEder Robles Morales100% (2)
- Dieta Fodmap HipocaloricaDocument5 pagesDieta Fodmap HipocaloricaMaría Patricia Jiménez Álvarez100% (1)
- Adicción Al Móvil o Celular - FolletoDocument2 pagesAdicción Al Móvil o Celular - FolletoEvelyn Karen Tito ChoqueNo ratings yet
- Enarm 2006 Cto 2Document34 pagesEnarm 2006 Cto 2mir-medicina100% (1)
- Cuáles Son Las Ventajas y Desventajas Que Tiene La Citogenética Molecular (Mateo Ruiz Florez)Document22 pagesCuáles Son Las Ventajas y Desventajas Que Tiene La Citogenética Molecular (Mateo Ruiz Florez)Bryan Stiven Bedoya100% (1)