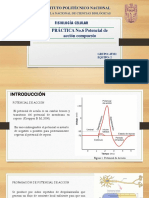
You might also like
- Microbiota RuminalDocument14 pagesMicrobiota RuminalCamila41No ratings yet
- Reprodução das cecílias: modos reprodutivos e cuidados parentaisDocument2 pagesReprodução das cecílias: modos reprodutivos e cuidados parentaisIonara NunesNo ratings yet
- Acidente com material biológicoDocument45 pagesAcidente com material biológicoRose FigueiredoNo ratings yet
- Anatomia Fisiologia BiologiaDocument140 pagesAnatomia Fisiologia BiologiaJéssica SeixasNo ratings yet
- Endocrinologia Oncologia Hematologia e Disturbios ImunorrelacionadosDocument156 pagesEndocrinologia Oncologia Hematologia e Disturbios ImunorrelacionadosAndré Barreto GuimarãesNo ratings yet
- Potencial de acción compuesto en nervio ciático de ranaDocument33 pagesPotencial de acción compuesto en nervio ciático de ranaPaoli RuizNo ratings yet
- Humanidade e Animalidade segundo Tim IngoldDocument2 pagesHumanidade e Animalidade segundo Tim IngoldMariana SimõesNo ratings yet
- Apostila Agronomia DemonstrativaDocument40 pagesApostila Agronomia DemonstrativaLuiz Fernandes100% (1)
- Origem da Vida - Processos IniciaisDocument10 pagesOrigem da Vida - Processos IniciaisHiago Leite da SilvaNo ratings yet
- Prova de biologia sobre células, fotossíntese e respiraçãoDocument3 pagesProva de biologia sobre células, fotossíntese e respiraçãoNathalia LopesNo ratings yet
- 1 Atividade 1 Ano 4 BimestreDocument2 pages1 Atividade 1 Ano 4 Bimestrevalfreefires21No ratings yet
- Sistema Imunitário SecundárioDocument5 pagesSistema Imunitário SecundárioMaria Maia100% (1)
- Câncer de Colo UteroDocument41 pagesCâncer de Colo UteroLuiza LimaNo ratings yet
- Características morfológicas e ciclos de vida de microrganismos parasitasDocument6 pagesCaracterísticas morfológicas e ciclos de vida de microrganismos parasitasRafael De AlmeidaNo ratings yet
- O que é Educação FísicaDocument23 pagesO que é Educação FísicaJuan Bueno100% (1)
- Resumo - Mitose e MeioseDocument2 pagesResumo - Mitose e MeioseIsadora TouricesNo ratings yet
- Cbo - Doenças Externas Oculares e Córnea 2013-14 PDFDocument616 pagesCbo - Doenças Externas Oculares e Córnea 2013-14 PDFLuana BragaNo ratings yet
- Ecossistemas e cadeias alimentaresDocument2 pagesEcossistemas e cadeias alimentarestati_lobatto100% (4)
- Site BiofísicaDocument8 pagesSite BiofísicaLuciclaudio Silva BarbosaNo ratings yet
- Exercícios - CiênciasDocument2 pagesExercícios - CiênciasMonica AlvesNo ratings yet
- Histologia - HemocipoieseDocument62 pagesHistologia - HemocipoieseMarcelle MarquesNo ratings yet
- Virus e Viroses - 3º Ano - Bio - 01Document58 pagesVirus e Viroses - 3º Ano - Bio - 01Igor CajatyNo ratings yet
- Sistema Nervoso MiotentericoDocument20 pagesSistema Nervoso MiotentericoJorge CamargoNo ratings yet
- Fisiopatologia e Dietoterapia Das Enfermidades Gástricas 2021.1Document115 pagesFisiopatologia e Dietoterapia Das Enfermidades Gástricas 2021.1Grazielle HugueninNo ratings yet
- Ecologia humana e sociabilidade urbanaDocument36 pagesEcologia humana e sociabilidade urbanadueduardaNo ratings yet
- Alimentacao SaudavelDocument22 pagesAlimentacao SaudavelGabriela RodriguesNo ratings yet
- Farmacocinética e formas de administração de medicamentosDocument59 pagesFarmacocinética e formas de administração de medicamentosnatalia100% (2)
- Avaliação de Ciências sobre Classificação de Seres Vivos e Evolução por Seleção NaturalDocument6 pagesAvaliação de Ciências sobre Classificação de Seres Vivos e Evolução por Seleção NaturalMônica CarvalhoNo ratings yet
- Microagulhamento Como Recurso de Tratamento NoDocument10 pagesMicroagulhamento Como Recurso de Tratamento NoIasmine PedrosoNo ratings yet
- Sociedade Primitiva PDFDocument20 pagesSociedade Primitiva PDFArquimedes AtlasNo ratings yet