You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
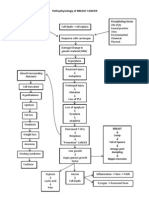
- Pathophysiology of BREAST CANCERDocument1 pagePathophysiology of BREAST CANCERAlinor Abubacar100% (6)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- DK Guide To The Human Body PDFDocument67 pagesDK Guide To The Human Body PDFTina Fishie Volf100% (5)
- ABX Pentra 60-C Plus Analyzer - Service ManualDocument314 pagesABX Pentra 60-C Plus Analyzer - Service ManualJose Rolando Orellana Rodriguez100% (2)
- Nci PBCF HTB22 - MCF7 - Sop 508 PDFDocument26 pagesNci PBCF HTB22 - MCF7 - Sop 508 PDFmumu4loveNo ratings yet
- Reaching and Being ReachedDocument18 pagesReaching and Being ReachedJoão Vitor Moreira Maia100% (1)
- U.S. Lessons Learned in Afghanistan: Hearing Committee On Foreign Affairs House of RepresentativesDocument112 pagesU.S. Lessons Learned in Afghanistan: Hearing Committee On Foreign Affairs House of Representativesgeraldi radityaNo ratings yet
- Virus Disinfection MethodDocument7 pagesVirus Disinfection Methodgeraldi radityaNo ratings yet
- Multikeyboard PDFDocument126 pagesMultikeyboard PDFgeraldi radityaNo ratings yet
- Disease and Disorder PhobiaDocument6 pagesDisease and Disorder Phobiageraldi radityaNo ratings yet
- F-16 Aircraft General Manual For Air Force StudiesDocument305 pagesF-16 Aircraft General Manual For Air Force StudiesAbdul Qadeer KhanNo ratings yet
- NARA GuidelinesDocument87 pagesNARA GuidelinesRoman VarasNo ratings yet
- Flexor Hallucis Longus Tendinitis-A Case of Mistaken Identity and Posterior Impingement Syndrome in Dancers: Evaluation and ManagementDocument3 pagesFlexor Hallucis Longus Tendinitis-A Case of Mistaken Identity and Posterior Impingement Syndrome in Dancers: Evaluation and Managementgeraldi radityaNo ratings yet
- Myocardial InfarctionDocument8 pagesMyocardial Infarctiongeraldi radityaNo ratings yet
- Current Concepts Review: Plantar FibromatosisDocument7 pagesCurrent Concepts Review: Plantar Fibromatosisgeraldi radityaNo ratings yet
- Bartybee Has A Go At: Building An Arduino 'Pro Micro' MPU-9150 Head TrackerDocument4 pagesBartybee Has A Go At: Building An Arduino 'Pro Micro' MPU-9150 Head Trackergeraldi radityaNo ratings yet
- Foot & Ankle InternationalDocument7 pagesFoot & Ankle Internationalgeraldi radityaNo ratings yet
- Foot & Ankle International: Current Concepts Review: Hallux RigidusDocument11 pagesFoot & Ankle International: Current Concepts Review: Hallux Rigidusgeraldi radityaNo ratings yet
- A Controlled Trial of Erenumab For Episodic MigraineDocument10 pagesA Controlled Trial of Erenumab For Episodic MigraineBryan Tam ArevaloNo ratings yet
- Osteomyelitis A Current ChangeDocument6 pagesOsteomyelitis A Current Changegeraldi radityaNo ratings yet
- Marina 2019Document4 pagesMarina 2019geraldi radityaNo ratings yet
- Cabeza y CuelloDocument35 pagesCabeza y CuelloLuis Álvaro Castillo CerveraNo ratings yet
- Chong2017 PDFDocument5 pagesChong2017 PDFgeraldi radityaNo ratings yet
- Viral Role in Sudden Hearing Loss ReviewDocument8 pagesViral Role in Sudden Hearing Loss Reviewgeraldi radityaNo ratings yet
- Decompressive Craniectomy: Indications and Techniques: Craniotomia Descompressiva: Indicações e TécnicasDocument6 pagesDecompressive Craniectomy: Indications and Techniques: Craniotomia Descompressiva: Indicações e Técnicasgeraldi radityaNo ratings yet
- (10920684 - Neurosurgical Focus) The Biophysical Role of Hemodynamics in The Pathogenesis of Cerebral Aneurysm Formation and RuptureDocument9 pages(10920684 - Neurosurgical Focus) The Biophysical Role of Hemodynamics in The Pathogenesis of Cerebral Aneurysm Formation and Rupturegeraldi radityaNo ratings yet
- JBI Critical Appraisal-Checklist For Analytical Cross Sectional StudiesDocument6 pagesJBI Critical Appraisal-Checklist For Analytical Cross Sectional StudieslinlinNo ratings yet
- Rupture of Lenticulostriate Artery AneurysmsDocument8 pagesRupture of Lenticulostriate Artery Aneurysmsgeraldi radityaNo ratings yet
- Dharma2015 Epid Stemi IndoDocument8 pagesDharma2015 Epid Stemi Indogeraldi radityaNo ratings yet
- Corsair CX750M ATX Power Supply - ManualDocument7 pagesCorsair CX750M ATX Power Supply - Manualcesinco0% (1)
- Basic Kes Zavoreo Demarin 355 359 PDFDocument5 pagesBasic Kes Zavoreo Demarin 355 359 PDFgeraldi radityaNo ratings yet
- GuifeDocument1 pageGuifegeraldi radityaNo ratings yet
- Css Prediktor StrokeDocument6 pagesCss Prediktor StrokeAnonymous l3X3jf0NPNo ratings yet
- Critical Appraisal For Cross SectionalDocument8 pagesCritical Appraisal For Cross SectionalSiti Zulaikhah100% (2)
- Working Hours Act Regulations for Employers and EmployeesDocument32 pagesWorking Hours Act Regulations for Employers and Employeesgeraldi radityaNo ratings yet
- Bajema2018 ISNRPSDocument8 pagesBajema2018 ISNRPSgeraldi radityaNo ratings yet
- CAFFEINEDocument2 pagesCAFFEINEVALDEZ, Teresita B.No ratings yet
- 7/23/2016 Cristina S. Nebres Mindanao State University at Naawan 1Document32 pages7/23/2016 Cristina S. Nebres Mindanao State University at Naawan 1Laila UbandoNo ratings yet
- The Science of Nei Jia Quan ExplainedDocument12 pagesThe Science of Nei Jia Quan ExplainedArran O RuaircNo ratings yet
- Burn OutDocument8 pagesBurn OutAtiqahAzizanNo ratings yet
- Pancreas - Pathological Practice and Research - K. Suda (Karger, 2007) WW PDFDocument329 pagesPancreas - Pathological Practice and Research - K. Suda (Karger, 2007) WW PDFIonut-Stefan CiobaneluNo ratings yet
- Essencial Phrasal VerbsDocument4 pagesEssencial Phrasal VerbsalvereNo ratings yet
- Fundamental PositionsDocument35 pagesFundamental PositionsVinod Kumar50% (4)
- Porges-2003-The Polyvagal Theory-Phylogenetic Contributions To Social BehaviorDocument11 pagesPorges-2003-The Polyvagal Theory-Phylogenetic Contributions To Social BehaviorJohn Bakalis100% (1)
- Biyani's Think Tank: Cell Biology & GeneticsDocument81 pagesBiyani's Think Tank: Cell Biology & GeneticsAkshay chandrakarNo ratings yet
- Ithomiini 2Document26 pagesIthomiini 2Roberta MirandaNo ratings yet
- Illusions The Magic Eye of PerceptionDocument12 pagesIllusions The Magic Eye of PerceptionArctic Illusion100% (2)
- ECG Localization of Culprit Artery in Acute Myocardial InfarctionDocument104 pagesECG Localization of Culprit Artery in Acute Myocardial Infarctionginaul100% (1)
- In This Issue: Wound Infection and ColonisationDocument7 pagesIn This Issue: Wound Infection and ColonisationSeftiana WahyuniNo ratings yet
- General Medicine and Surgery For Dental Practitioners - Part 1. History Taking and Examination of The Clothed PatientDocument4 pagesGeneral Medicine and Surgery For Dental Practitioners - Part 1. History Taking and Examination of The Clothed Patientanees jamalNo ratings yet
- ABCs of CPR: A Simple Life-Saving ProcedureDocument3 pagesABCs of CPR: A Simple Life-Saving ProcedureChanthini VinayagamNo ratings yet
- Pharmaceutics 09 00041 v2Document14 pagesPharmaceutics 09 00041 v2thasyaNo ratings yet
- Quiz ReviewerDocument8 pagesQuiz ReviewerCai PascualNo ratings yet
- Nontoxic Nodular GoiterDocument7 pagesNontoxic Nodular GoiterKayshey Christine ChuaNo ratings yet
- Circulatory System - Class 10Document4 pagesCirculatory System - Class 10Biozeneca classesNo ratings yet
- Blood ? Summary PDFDocument10 pagesBlood ? Summary PDFmanar aleneziNo ratings yet
- Safe IV CannulationDocument32 pagesSafe IV Cannulationanon_281166672No ratings yet
- BLOCK II LMS Quiz AnatomyDocument27 pagesBLOCK II LMS Quiz AnatomyAshley BuchananNo ratings yet
- Test Bank For Anatomy and Physiology 10th Edition by PattonDocument32 pagesTest Bank For Anatomy and Physiology 10th Edition by PattonCassandraDuncanmcytd100% (34)
- Chapman System of ClassificationDocument6 pagesChapman System of Classificationvineetvishal73No ratings yet
- Cysteine, Methionine, ProlineDocument3 pagesCysteine, Methionine, ProlineRio BurlazaNo ratings yet