You might also like
- Biología 1 Guía de Aprendizaje de U3 - S1Document10 pagesBiología 1 Guía de Aprendizaje de U3 - S1alajoapoNo ratings yet
- Guía EmbriologíaDocument9 pagesGuía EmbriologíaJennifer Yucra CamposanoNo ratings yet
- Desarrollo embrionario inicial en aves y mamíferosDocument9 pagesDesarrollo embrionario inicial en aves y mamíferosjose luis sotoNo ratings yet
- Guía Académica Biología Noveno Ii Per. - 2021Document10 pagesGuía Académica Biología Noveno Ii Per. - 2021LUZ AVILEZNo ratings yet
- Modulo AvanzadoDocument50 pagesModulo AvanzadoJhons Jara SamameNo ratings yet
- U3 Actividad10 MCHyBDocument19 pagesU3 Actividad10 MCHyBJorge TamezNo ratings yet
- 5CN G7 Como Se Clasifican de OrganismosDocument7 pages5CN G7 Como Se Clasifican de Organismoswilson de arcoNo ratings yet
- Biologia Primero MedioDocument2 pagesBiologia Primero MedioIsidora LagosNo ratings yet
- BIOQUMICADocument5 pagesBIOQUMICAROOSEVELT AUGUSTO TELLO REYNANo ratings yet
- Biologia FebreroDocument15 pagesBiologia FebreroTodoTesis Chaguaramas2023No ratings yet
- 1.feria012 01 La Intimidad Del Gorgojo Chino Al DescubiertoDocument22 pages1.feria012 01 La Intimidad Del Gorgojo Chino Al DescubiertoAnonymous yR8Hgcb3yNo ratings yet
- Tarea 0 - 7554 - Fundamentos Sobre Fisiologia Animal - AmaguaDocument6 pagesTarea 0 - 7554 - Fundamentos Sobre Fisiologia Animal - AmaguaAMAGUA SANGOQUIZA JEFFERSON VINICIONo ratings yet
- La nueva clasificación de los gasterópodosDocument9 pagesLa nueva clasificación de los gasterópodosARIANA YOLANDA CERIN ECHENo ratings yet
- La-Evolución-para-Sexto-de-Primaria 2Document4 pagesLa-Evolución-para-Sexto-de-Primaria 2SANDRA MILENA AYALA LENISNo ratings yet
- Examen Previo 2do-3 - 231107 - 143443Document9 pagesExamen Previo 2do-3 - 231107 - 143443Abril SivesNo ratings yet
- La Evolución para Sexto de PrimariaDocument4 pagesLa Evolución para Sexto de PrimariaJulieth Camej100% (2)
- La Evolución para Sexto de PrimariaDocument4 pagesLa Evolución para Sexto de PrimariaYuli Ore Huamalias50% (2)
- Evolucion de Los Ojos y FotorreceptoresDocument6 pagesEvolucion de Los Ojos y FotorreceptoresMauricio MoralesNo ratings yet
- Dialnet DinamicaFolicularOvaricaEnVacasRepetidoras 1209196Document12 pagesDialnet DinamicaFolicularOvaricaEnVacasRepetidoras 1209196lauraNo ratings yet
- La Evolución para Sexto Grado de PrimariaDocument4 pagesLa Evolución para Sexto Grado de PrimariaJoan Diaz AliagaNo ratings yet
- Actividades indagar evolución especiaciónDocument2 pagesActividades indagar evolución especiaciónWilber García RamirezNo ratings yet
- Biologia Segundo Medio Clase 4 Teorias EvolutivasDocument16 pagesBiologia Segundo Medio Clase 4 Teorias EvolutivasDominique bucareyNo ratings yet
- Parte de Monografia de Medicina LegalDocument5 pagesParte de Monografia de Medicina LegalXiomara Alfaro AraujoNo ratings yet
- La clasificación de organismos y la biodiversidadDocument10 pagesLa clasificación de organismos y la biodiversidadLuis Angel Gárate ChambillaNo ratings yet
- CJRG - Practica Analisis en FrescoDocument6 pagesCJRG - Practica Analisis en FrescoLupita Ramos GracianoNo ratings yet
- GUIAS DE APRENDIZAJE #3 Oscar Garcia GRADO NOVENO Ciencias Naturales-ArtisticaDocument12 pagesGUIAS DE APRENDIZAJE #3 Oscar Garcia GRADO NOVENO Ciencias Naturales-ArtisticaOscarAugustoGarciaAlvarezNo ratings yet
- Informe Anatomia de PecesDocument2 pagesInforme Anatomia de PecesHangarNo ratings yet
- Tema#1 La Evolución de Las Especies X °Document10 pagesTema#1 La Evolución de Las Especies X °Abilio MontenegroNo ratings yet
- Aguirre 2015Document9 pagesAguirre 2015Arturo VillanuevaNo ratings yet
- Evolución biológica: Teoría sintética, adaptación y cambios en especiesDocument3 pagesEvolución biológica: Teoría sintética, adaptación y cambios en especiesMARCO ANTONIO ZUNIGA GONZALESNo ratings yet
- Ciclo Reproductivo de Cerion Mumia Chrysalis (Pulmonata StylommatophoraDocument7 pagesCiclo Reproductivo de Cerion Mumia Chrysalis (Pulmonata StylommatophoraAlexis SuarezNo ratings yet
- 19 1888 Sec Club de Revistas La Transparencia Como EstrategiaDocument3 pages19 1888 Sec Club de Revistas La Transparencia Como EstrategiaPestos SsNo ratings yet
- Organiz Cuerpo Eval Sol Prueba CompetencialDocument12 pagesOrganiz Cuerpo Eval Sol Prueba CompetenciallourdesNo ratings yet
- Diversidad y clasificación de animales vertebradosDocument8 pagesDiversidad y clasificación de animales vertebradosMagalí ScialfaNo ratings yet
- 90597Document12 pages90597Katy VillanuevaNo ratings yet
- Trabajo BiotecnologiaDocument24 pagesTrabajo BiotecnologiasoruizNo ratings yet
- Guia de Biologia de MayoDocument8 pagesGuia de Biologia de MayoMariana martinezNo ratings yet
- Guia para Segunda Tarea Exaula Ecologia y Ambiente 2022Document12 pagesGuia para Segunda Tarea Exaula Ecologia y Ambiente 2022Denisse Marisol Lemus VasquezNo ratings yet
- 2do - Anatomía y Fisiología AnimalDocument49 pages2do - Anatomía y Fisiología AnimalExequielPiacenzaNo ratings yet
- Dieta de AtheneDocument9 pagesDieta de AtheneGénesis EscobarNo ratings yet
- Pae (1) - MV (A) - Etologia - Teoria Del Etograma Pata CriaDocument16 pagesPae (1) - MV (A) - Etologia - Teoria Del Etograma Pata Criaandres09coronelNo ratings yet
- 4°-Clasificación SV-Estudiantes - 230927 - 195404Document16 pages4°-Clasificación SV-Estudiantes - 230927 - 195404Ines Beatriz TrebeNo ratings yet
- Content ServerDocument11 pagesContent Serverbrisa de los angeles zamoraperezNo ratings yet
- MurciélagosDocument17 pagesMurciélagosGerman Andres Lavin SouterNo ratings yet
- La taxonomía de LinneoDocument9 pagesLa taxonomía de LinneoivanNo ratings yet
- Tomo I Libro Biodiversidad Chile MMA BioExtintaDocument16 pagesTomo I Libro Biodiversidad Chile MMA BioExtintaJhoann CantoNo ratings yet
- Cerebro EvolucionDocument10 pagesCerebro Evolucionapi-371953967% (3)
- 1era Semana BIOLOGIA - GENERALIDADESDocument53 pages1era Semana BIOLOGIA - GENERALIDADESLuisNo ratings yet
- Bianchi Evolucion PDFDocument12 pagesBianchi Evolucion PDFSylvia Stella G. TroncosoNo ratings yet
- Anestesia Y Analgesia en Reptiles - Selecciones VeterinariasDocument38 pagesAnestesia Y Analgesia en Reptiles - Selecciones VeterinariasandresoliverosgomezNo ratings yet
- Datos Curiosos Del TiburónDocument7 pagesDatos Curiosos Del TiburónNAYELI RAMIREZNo ratings yet
- Métodos para estudiar avesDocument4 pagesMétodos para estudiar avesNoticiero Escolar AbsNo ratings yet
- Evaluación de Recursos HidrobiologicosDocument21 pagesEvaluación de Recursos HidrobiologicosDiego AcuñaNo ratings yet
- Observación y Reconocimiento de ProtozoosDocument16 pagesObservación y Reconocimiento de ProtozoostatianaNo ratings yet
- ACTIVIDADES RECUPERACIÓN 1ºESO TEMAS 5 Y 6 Biosfera y Animales VertebradosDocument8 pagesACTIVIDADES RECUPERACIÓN 1ºESO TEMAS 5 Y 6 Biosfera y Animales VertebradosGuillermo GPNo ratings yet
- Capítulo 3 Zoología Primera ParteDocument60 pagesCapítulo 3 Zoología Primera ParteErick AlvarezNo ratings yet
- Manejo de Tyto Alba en ZoologicoDocument3 pagesManejo de Tyto Alba en ZoologicoVictor Velasquez Zea100% (1)
- La clasificación de los seres vivos en cinco reinosDocument17 pagesLa clasificación de los seres vivos en cinco reinosHENRY RODRIGUEZNo ratings yet
- Aves Marinas y Playeras de ChileDocument2 pagesAves Marinas y Playeras de ChileXimena Olivares CastilloNo ratings yet
- Aves Del Rio UruguayDocument254 pagesAves Del Rio UruguayhernangenNo ratings yet
- BorradorVeterinaria PINAR PDFDocument39 pagesBorradorVeterinaria PINAR PDFXimena Olivares CastilloNo ratings yet
- 1° Evaluación3 Mayo Letras M, L, P, S, DDocument4 pages1° Evaluación3 Mayo Letras M, L, P, S, DXimena Olivares CastilloNo ratings yet
- 1° Evaluación4 Junio Letras M, L, P, S, D, Ca, Que Qui, Co, Cu, NDocument4 pages1° Evaluación4 Junio Letras M, L, P, S, D, Ca, Que Qui, Co, Cu, NXimena Olivares CastilloNo ratings yet
- Los Humedales Cuidan El Agua - Unesco PDFDocument16 pagesLos Humedales Cuidan El Agua - Unesco PDFmiguelvelardeNo ratings yet
- 2° Evaluación1 Lectura Comprensiva AbrilDocument5 pages2° Evaluación1 Lectura Comprensiva AbrilEliana Castillo D.No ratings yet
- AnovaDocument13 pagesAnovaNicolas BrunieNo ratings yet
- El Riñón Del Gato 1Document145 pagesEl Riñón Del Gato 1Ximena Olivares Castillo100% (1)
- Ecología Clase 7-8Document27 pagesEcología Clase 7-8Ximena Olivares CastilloNo ratings yet
- 1° Evaluación2 Abril Letras M, L, PDocument6 pages1° Evaluación2 Abril Letras M, L, PXimena Olivares CastilloNo ratings yet
- Condor Monografia PDFDocument8 pagesCondor Monografia PDFyoselin ingretel villanueva muñoz100% (2)
- Conservación Aves Playeras PacíficoDocument88 pagesConservación Aves Playeras PacíficoXimena Olivares Castillo100% (1)
- Monte Patria 27 de Julio PDFDocument2 pagesMonte Patria 27 de Julio PDFXimena Olivares CastilloNo ratings yet
- Impacto de La Contaminación Lumínica Sobre Las AvesDocument11 pagesImpacto de La Contaminación Lumínica Sobre Las AvesXimena Olivares CastilloNo ratings yet
- Manual de quesos y lácteosDocument48 pagesManual de quesos y lácteoscerda69No ratings yet
- Revista Veteros No1 2016Document41 pagesRevista Veteros No1 2016Ximena Olivares CastilloNo ratings yet
- Flora Acuática y Palustre Introducida en ChileDocument94 pagesFlora Acuática y Palustre Introducida en ChileXimena Olivares CastilloNo ratings yet
- Manejo de Fracturas en AvesDocument13 pagesManejo de Fracturas en AvesGabriel Di GennaroNo ratings yet
- 04 Fluidoterpia PracticaDocument8 pages04 Fluidoterpia PracticajaeboomNo ratings yet
- Hipoadrenocorticismo - en - Perros - Vitoria - 2014 PDFDocument6 pagesHipoadrenocorticismo - en - Perros - Vitoria - 2014 PDFJorge VergaraNo ratings yet
- 50 255 3 PBDocument14 pages50 255 3 PBSandra Alicia RomeroNo ratings yet
- Herpes Virus Bovino y Diarrea Viral Bovina PDFDocument8 pagesHerpes Virus Bovino y Diarrea Viral Bovina PDFXimena Olivares CastilloNo ratings yet
- Seminario 2 Ectima ContagiosoDocument17 pagesSeminario 2 Ectima ContagiosoXimena Olivares CastilloNo ratings yet
- Influencia Aviar Produccion IIDocument36 pagesInfluencia Aviar Produccion IIXimena Olivares CastilloNo ratings yet
- Alimentacion Del Paciente SilvestreDocument9 pagesAlimentacion Del Paciente SilvestreXimena Olivares CastilloNo ratings yet
- Análisis del cuento El almohadón de plumas de Horacio QuirogaDocument9 pagesAnálisis del cuento El almohadón de plumas de Horacio QuirogaXimena Olivares CastilloNo ratings yet
- La Maquina Del Tiempo GuíaDocument14 pagesLa Maquina Del Tiempo Guíamalrogo252No ratings yet
- Defensas RibereñasDocument12 pagesDefensas RibereñasRyduan Cristophers Sara Gutierrez100% (1)
- HERAZO REYES DANIEL NeumoníaDocument21 pagesHERAZO REYES DANIEL NeumoníaDaniel HerazoNo ratings yet
- Productos Rabago 2016 R1, R2 y R8 TalentuaDocument39 pagesProductos Rabago 2016 R1, R2 y R8 TalentuaCristie MtzNo ratings yet
- Métodos de EsterilizaciónDocument6 pagesMétodos de EsterilizaciónEdith Lopez Gomez100% (1)
- Teg Ortopedia .Botello AnteDocument70 pagesTeg Ortopedia .Botello AnteAlexis LeonNo ratings yet
- Las Defensas Del Organismo HumanoDocument10 pagesLas Defensas Del Organismo HumanoMili GarcíaNo ratings yet
- PARASHA 14 VAER - para Ni - Os de 6-10 A - OsDocument10 pagesPARASHA 14 VAER - para Ni - Os de 6-10 A - OsMario KoganNo ratings yet
- Analisis de Factibilidad Financiera de Construccion de Torre de ApartamentosDocument2 pagesAnalisis de Factibilidad Financiera de Construccion de Torre de ApartamentosCarmen ArizaNo ratings yet
- Manual de Procedimientos Operativos Estándar para El Centro de Rehabilitación de Fauna Silvestre de Victoria, CaldasDocument38 pagesManual de Procedimientos Operativos Estándar para El Centro de Rehabilitación de Fauna Silvestre de Victoria, CaldasJulia Maria Ribeiro100% (1)
- ANTIMICOBACTERIANOSDocument30 pagesANTIMICOBACTERIANOSapi-3750373100% (1)
- Recibo Movistar Julio 2021Document4 pagesRecibo Movistar Julio 2021Amadeo Peralta RodriguezNo ratings yet
- Manual de procesos de evaluación para la construcción de cubierta en estructura metálicaDocument2 pagesManual de procesos de evaluación para la construcción de cubierta en estructura metálicasoniaNo ratings yet
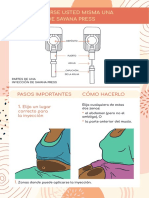
- Instrucciones Autoinyeccion Sayana PressDocument5 pagesInstrucciones Autoinyeccion Sayana PressNeyda GuevaraNo ratings yet
- Electrorefinación de Ánodos de PlomoDocument2 pagesElectrorefinación de Ánodos de PlomoAngel Luis Uscuchagua PomaNo ratings yet
- Razonamiento Verbal Solucionario 07: Ceprunsa 2021 Ii FaseDocument8 pagesRazonamiento Verbal Solucionario 07: Ceprunsa 2021 Ii FaseAlahi UscaNo ratings yet
- Grout MorteroDocument2 pagesGrout MorteroRichard GarciaNo ratings yet
- Analisis Capitulo 5 de Psicólogia y MasasDocument7 pagesAnalisis Capitulo 5 de Psicólogia y MasasMike T. FrancoNo ratings yet
- RosáceaDocument5 pagesRosáceaSofia Vela PNo ratings yet
- Monografia PDFDocument81 pagesMonografia PDFzulyleonardoNo ratings yet
- Revista+2020 2021+ +edit +ort 713 723Document11 pagesRevista+2020 2021+ +edit +ort 713 723Miguel FigueroaNo ratings yet
- Promoción de la salud ocupacionalDocument9 pagesPromoción de la salud ocupacionalgonzalo vasquezNo ratings yet
- Cocmoelec Pro0 Op 03 de Soldadura ExotermicaDocument9 pagesCocmoelec Pro0 Op 03 de Soldadura ExotermicaNelsonNo ratings yet
- Rinoplastia: Cirugía de corrección y perfeccionamiento de la narizDocument52 pagesRinoplastia: Cirugía de corrección y perfeccionamiento de la narizSonia BermudezNo ratings yet
- 02 Cronograma de Adquisicion de MaterialesDocument9 pages02 Cronograma de Adquisicion de MaterialesRichard HuamánNo ratings yet
- Hematologia ForenseDocument35 pagesHematologia ForenseSammael GamonalNo ratings yet
- Programa de Desarrollo PsicomotorDocument8 pagesPrograma de Desarrollo PsicomotorEduardo Cristofer Insuasti Ortega50% (2)
- Ruta de Abusexual Definitiva 2008Document3 pagesRuta de Abusexual Definitiva 2008sebastianNo ratings yet
- La Muerte Del FaloDocument11 pagesLa Muerte Del FaloGERMAN BELTRANNo ratings yet
- Carbonato de CalcioDocument2 pagesCarbonato de CalcioRosmery BelloNo ratings yet
- Manual SGADOR 24x17 - WEB - 20 03Document5 pagesManual SGADOR 24x17 - WEB - 20 03CARLOS DANIEL DUARTE ZACAPALANo ratings yet