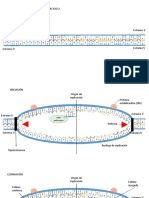
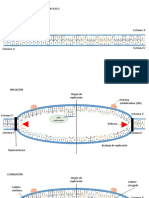
You might also like
- Citoesqueleto ForoDocument39 pagesCitoesqueleto ForoClaudia Montes GuerraNo ratings yet
- Informe BioinformaticaDocument15 pagesInforme Bioinformaticadiana medinaNo ratings yet
- EnzimasDocument39 pagesEnzimasAbel ForesNo ratings yet
- Metabolismo Del NitrogenoDocument36 pagesMetabolismo Del NitrogenoGeovanni moralesNo ratings yet
- Hidrolisis Del Hipurato Sódico.Document8 pagesHidrolisis Del Hipurato Sódico.Adrian Espinoza PalominoNo ratings yet
- Cuestionario 11Document2 pagesCuestionario 11Mayker PanibraNo ratings yet
- 1208 Es 1 PDFDocument2 pages1208 Es 1 PDFWendy Iveth Avelar AcostaNo ratings yet
- Lab 2. Extracción y Cuantificacion Del ADNDocument4 pagesLab 2. Extracción y Cuantificacion Del ADNEliana Rocio Moya Bare�oNo ratings yet
- Dna PlasmidicoDocument2 pagesDna PlasmidicojorgeNo ratings yet
- Re Sum enDocument7 pagesRe Sum enJD ChaverriNo ratings yet
- Membrana celular: estructura y funciones enDocument37 pagesMembrana celular: estructura y funciones enErika ToponNo ratings yet
- Resumen Ciclo de CoriDocument10 pagesResumen Ciclo de CoriMarisa LeyvaNo ratings yet
- Informe de ADN en La CebollaDocument7 pagesInforme de ADN en La CebollaANGELA NICOLE REINA BARBOSANo ratings yet
- Factores Que Inhiben La MitosisDocument3 pagesFactores Que Inhiben La MitosisDavis AvilbarNo ratings yet
- Practica 3 y 4 Diversidad CelularDocument4 pagesPractica 3 y 4 Diversidad CelularmarulasNo ratings yet
- Manejo y vías de administración en el ratón de laboratorioDocument40 pagesManejo y vías de administración en el ratón de laboratorioSofi CortesNo ratings yet
- Especialización de La MembranaDocument2 pagesEspecialización de La MembranaMarisol100% (2)
- GENETICADocument25 pagesGENETICAJhamil LopezNo ratings yet
- HISTOLOGIADocument7 pagesHISTOLOGIAMaría Valentina Rubiano PeñaNo ratings yet
- Práctica 7 Actividad Enzimática de Las Peroxidasas.Document6 pagesPráctica 7 Actividad Enzimática de Las Peroxidasas.Agustin Pedraza GaleanaNo ratings yet
- Practica #5.DETERMINACION DE LINFOCITOS T Y B.Document9 pagesPractica #5.DETERMINACION DE LINFOCITOS T Y B.JORGE EMILIANO RAMOS VILLALOBOS100% (1)
- Cuál Es La Diferencia Entre Célula Animal y VegetalDocument27 pagesCuál Es La Diferencia Entre Célula Animal y VegetalANGELANo ratings yet
- BiologíaDocument38 pagesBiologíabunny EspejoNo ratings yet
- Guia 5 - Permeabilidad Celular - ERITROCITOS Y ELODEADocument8 pagesGuia 5 - Permeabilidad Celular - ERITROCITOS Y ELODEANIKOLE SANABRIA RODRIGUEZNo ratings yet
- Taller Clasificación de Enzimas II 2020Document4 pagesTaller Clasificación de Enzimas II 2020catailnaNo ratings yet
- Determinación de proteínas mediante pruebas químicasDocument7 pagesDeterminación de proteínas mediante pruebas químicasNathalie DávilaNo ratings yet
- Infografia ProtozoariosDocument1 pageInfografia ProtozoariosChrystal CampilloNo ratings yet
- Reporte Practica 1Document9 pagesReporte Practica 1Carlos Cerón LinaresNo ratings yet
- Ensayo BuffersDocument4 pagesEnsayo Buffersleslie cadenillasNo ratings yet
- Hidrolizis de IpidosDocument7 pagesHidrolizis de IpidosJorge ContrerasNo ratings yet
- Proceso de Maduracion de Los Diferentes ArnDocument4 pagesProceso de Maduracion de Los Diferentes ArnJosselynSánchezYRichardBarrezuetaNo ratings yet
- E-2 y E-3 Cuadro Sinoptico Biomoleculas PDFDocument1 pageE-2 y E-3 Cuadro Sinoptico Biomoleculas PDFRaul Ieled Catan IafeNo ratings yet
- Taller de Bioquimica ProteinaDocument7 pagesTaller de Bioquimica ProteinaRosman Ramos MartinezNo ratings yet
- Mecanismos de Regulación EnzimáticaDocument2 pagesMecanismos de Regulación EnzimáticaRobert Isaias Mendoza CardenasNo ratings yet
- Practica 1 HelmintosDocument6 pagesPractica 1 Helmintosdev3mondrag3n100% (1)
- Fosforilación OxidativaDocument2 pagesFosforilación OxidativaKarryMuñizLauraNo ratings yet
- PRÁCTICA #08 EnzimasDocument6 pagesPRÁCTICA #08 Enzimasjimmy gomezNo ratings yet
- Cuestionario Práctica 6 - Técnicas de Tinción DiferencialesDocument4 pagesCuestionario Práctica 6 - Técnicas de Tinción DiferencialesLizzet SalazarNo ratings yet
- Aislamiento de ADN PlasmídicoDocument2 pagesAislamiento de ADN PlasmídicoZuri ManriqueNo ratings yet
- Cuadro Comparativo Tejido ConectivoDocument1 pageCuadro Comparativo Tejido ConectivoDulce FloresNo ratings yet
- 1 - MetazoariosDocument34 pages1 - Metazoariostara100% (1)
- Informe 10 - Hidrólisis de LípidosDocument9 pagesInforme 10 - Hidrólisis de LípidosDiana GrimaldoNo ratings yet
- Extracción y Cuantificación de Ácidos NucleicosDocument6 pagesExtracción y Cuantificación de Ácidos NucleicosLucero Odar CastilloNo ratings yet
- CitoesqueletoDocument50 pagesCitoesqueletoBrian FariñaNo ratings yet
- Manual Práctico de Imagenología en Grandes AnimalesDocument30 pagesManual Práctico de Imagenología en Grandes AnimalesClaudia CuitivaNo ratings yet
- Estudios ToxicológicosDocument17 pagesEstudios ToxicológicosleoNo ratings yet
- Biologia 1 PDFDocument4 pagesBiologia 1 PDFKathe SierraNo ratings yet
- GlosarioDocument5 pagesGlosariojavier elias salcedo ramosNo ratings yet
- Control de Expresión GénicaDocument43 pagesControl de Expresión GénicaDIANA CAROLINA DIAZ JIMENEZ91% (11)
- 2do-Informe-De-Genetica 06-09-2017Document11 pages2do-Informe-De-Genetica 06-09-2017edmundoNo ratings yet
- Laboratorio-Técnicas de Tinción2Document5 pagesLaboratorio-Técnicas de Tinción2camilaleguizamon123No ratings yet
- Diversidad BacteriaDocument115 pagesDiversidad BacteriaSuarez GOmez Alexis Gabriel100% (1)
- Práctica 5 - Identificación de HongosDocument6 pagesPráctica 5 - Identificación de HongosMarcia OchoaNo ratings yet
- Taller 2 de Biología CelularDocument12 pagesTaller 2 de Biología Celularjhoanna aguirreNo ratings yet
- Atp BiologiaDocument2 pagesAtp BiologiaEl chirrete Del salónNo ratings yet
- Extracción ADN vegetalDocument9 pagesExtracción ADN vegetalAngela Salinas100% (1)
- Informe de FermentacionDocument24 pagesInforme de FermentacionSARANo ratings yet
- SINTROFIADocument14 pagesSINTROFIALeidy BaronaNo ratings yet
- La evolución del citoesqueleto: desde lo simple en procariotas a lo complejo en eucariotasDocument18 pagesLa evolución del citoesqueleto: desde lo simple en procariotas a lo complejo en eucariotasBrayan RodriguezNo ratings yet
- Evolucion Del CitoesqueletoDocument9 pagesEvolucion Del CitoesqueletoJhony Vargas ReyesNo ratings yet
- Clase 12 RsptasDocument2 pagesClase 12 RsptasalfredoNo ratings yet
- Adn ReDocument4 pagesAdn RealfredoNo ratings yet
- 1ro - Plan Lector - 1ra Actividad de DiciembreDocument1 page1ro - Plan Lector - 1ra Actividad de DiciembrealfredoNo ratings yet
- 1ro - Plan Lector - 1ra Actividad de DiciembreDocument1 page1ro - Plan Lector - 1ra Actividad de DiciembrealfredoNo ratings yet
- Helicobacter Pylori: y Enfermedades RelacionadasDocument5 pagesHelicobacter Pylori: y Enfermedades RelacionadasalfredoNo ratings yet
- EjerciciosDocument2 pagesEjerciciosalfredoNo ratings yet
- Clase 12Document40 pagesClase 12alfredoNo ratings yet
- Adn ReDocument4 pagesAdn RealfredoNo ratings yet
- Clase 12 RsptasDocument2 pagesClase 12 RsptasalfredoNo ratings yet
- Clase 9. Naturaleza Del Gen y El Genoma 2020-2Document24 pagesClase 9. Naturaleza Del Gen y El Genoma 2020-2alfredoNo ratings yet
- Adn ReDocument4 pagesAdn RealfredoNo ratings yet
- UNIDAD 5 Autonomía y Responsabilidad IndividualDocument3 pagesUNIDAD 5 Autonomía y Responsabilidad IndividualJocelyn Aranda RabanalNo ratings yet
- La Sexualidad AnaliticaDocument1 pageLa Sexualidad AnaliticaalfredoNo ratings yet
- 1 PDFDocument10 pages1 PDFalfredoNo ratings yet
- HHDocument8 pagesHHalfredoNo ratings yet
- Articulo de OpinionDocument5 pagesArticulo de OpinionalfredoNo ratings yet
- Polimorfismo de MHC-IIDocument1 pagePolimorfismo de MHC-IIalfredoNo ratings yet
- FiorellaDocument7 pagesFiorellaalfredoNo ratings yet
- Plan de Trabajo de FilosofiaDocument5 pagesPlan de Trabajo de FilosofiaalfredoNo ratings yet
- Guerra ADocument2 pagesGuerra AalfredoNo ratings yet
- Embarazo incipiente 5 semanas sin latido cardiacoDocument3 pagesEmbarazo incipiente 5 semanas sin latido cardiacoalfredoNo ratings yet
- MurrayDocument1 pageMurrayalfredoNo ratings yet
- Analisis CompetititvoDocument2 pagesAnalisis CompetititvoalfredoNo ratings yet
- Ensayo FinalDocument11 pagesEnsayo FinalalfredoNo ratings yet
- Contaminación Del Parque AutomotorDocument29 pagesContaminación Del Parque Automotorwillgeiner100% (5)
- Informe EstadisticoDocument30 pagesInforme EstadisticoalfredoNo ratings yet
- Contaminación Del Parque AutomotorDocument29 pagesContaminación Del Parque Automotorwillgeiner100% (5)
- Informe Estadistico TerminadoDocument31 pagesInforme Estadistico TerminadoalfredoNo ratings yet
- Informe Estadistico TerminadoDocument31 pagesInforme Estadistico TerminadoalfredoNo ratings yet
- Informe Estadistico TerminadoDocument31 pagesInforme Estadistico TerminadoalfredoNo ratings yet
- Planteamiento Del ProblemaDocument3 pagesPlanteamiento Del ProblemaPaola HernandezNo ratings yet
- Informe El TrebolDocument4 pagesInforme El TrebolAnthonyNo ratings yet
- Mendoza Sumoso Rodrigo Estudio Planta AnexosDocument96 pagesMendoza Sumoso Rodrigo Estudio Planta AnexosDaniel Torrico LupeNo ratings yet
- Oscar Jorge Bernal Luna: No. de ServicioDocument1 pageOscar Jorge Bernal Luna: No. de ServicioJesus TalamantesNo ratings yet
- Spsu-865 Ejercicio T001.Document12 pagesSpsu-865 Ejercicio T001.Valeria Prieto YsmiñoNo ratings yet
- Codigo de Etica Negocios InternacionalesDocument7 pagesCodigo de Etica Negocios InternacionalesDaniel MenjivarNo ratings yet
- Parques Eólicos MarinosDocument4 pagesParques Eólicos MarinosJhosep Rober pmNo ratings yet
- Solicitud de inscripción a mentorías empresarialesDocument2 pagesSolicitud de inscripción a mentorías empresarialesnelson ferroNo ratings yet
- Matriz Condiciones de SaludDocument28 pagesMatriz Condiciones de SaludJorgeNo ratings yet
- Inhibidor EnzimáticoDocument18 pagesInhibidor EnzimáticoAdri CruzNo ratings yet
- Optimización de atención al cliente en supermercados mediante pantallas táctilesDocument23 pagesOptimización de atención al cliente en supermercados mediante pantallas táctilesMarcos RojasNo ratings yet
- Programa y régimen de regularidad de Matemática 1Document4 pagesPrograma y régimen de regularidad de Matemática 1Carabajal LuciaNo ratings yet
- Borras Et Al - Acaparamiento de Tierras y Acumulación Capitalista. ASpectos Clave en América Latina PDFDocument29 pagesBorras Et Al - Acaparamiento de Tierras y Acumulación Capitalista. ASpectos Clave en América Latina PDFDiego Palacios LlaqueNo ratings yet
- Informe Maquinas 1 Marco TeoricoDocument7 pagesInforme Maquinas 1 Marco TeoricoAnonymous GcNo6ivNo ratings yet
- Más de 100 Ejemplos de Sustantivos PropiosDocument7 pagesMás de 100 Ejemplos de Sustantivos PropiosLiz Karina Aguirre100% (2)
- Catalogo Baterías Solares - ITHURBIDE - ISECOMDocument5 pagesCatalogo Baterías Solares - ITHURBIDE - ISECOMmcombaleNo ratings yet
- Actividad 4Document8 pagesActividad 4Ximena AlejandraNo ratings yet
- Entrenamiento de Karate ShotokanDocument2 pagesEntrenamiento de Karate ShotokanHctor AlvaradoNo ratings yet
- Sesión #1 Conceptos GeneralesDocument109 pagesSesión #1 Conceptos GeneralesJorge Javier Granda GranadinoNo ratings yet
- En Torno A La Prensa GraficaDocument35 pagesEn Torno A La Prensa GraficaVictoria PedroNo ratings yet
- Eca1 Calculo IDocument12 pagesEca1 Calculo IJosé Luis Cruz JiménezNo ratings yet
- PWS 2.3 Genx Spanish - R06 (07.2016)Document2 pagesPWS 2.3 Genx Spanish - R06 (07.2016)Christian Javier Lopez OrduzNo ratings yet
- DASH Versión ChilenaDocument3 pagesDASH Versión ChilenaDani Orellana MedinaNo ratings yet
- Deficit Fiscal EconomiaDocument5 pagesDeficit Fiscal EconomiaDavid Carvajal SánchezNo ratings yet
- Acueducto Paralelo Chicbul-CarmenDocument356 pagesAcueducto Paralelo Chicbul-CarmenmadiduzNo ratings yet
- Ejercicios de senoides y funciones forzadasDocument5 pagesEjercicios de senoides y funciones forzadasEdwin Mayares LópezNo ratings yet
- Signos VitalwsDocument21 pagesSignos VitalwsMaeva OlveraNo ratings yet
- DOC20190115145758Proyecto Completo Firmado PDFDocument413 pagesDOC20190115145758Proyecto Completo Firmado PDFTAMIFUENTESNo ratings yet
- A#3 AvcDocument7 pagesA#3 AvcAlejandra Vázquez CastejónNo ratings yet
- Certificado de Pozo A Tierra (Av. Prolongación Tacna 117) Rimac 09122019Document2 pagesCertificado de Pozo A Tierra (Av. Prolongación Tacna 117) Rimac 09122019Eduardo Garagundo HuancaNo ratings yet