You might also like
- MS 255 Assignment1Document3 pagesMS 255 Assignment1Rhaiza PabelloNo ratings yet
- Cancers 13 01521 v2Document20 pagesCancers 13 01521 v2Rhaiza PabelloNo ratings yet
- CancersDocument13 pagesCancersRhaiza PabelloNo ratings yet
- 1 - CA A Cancer Journal For CliniciansDocument6 pages1 - CA A Cancer Journal For CliniciansRhaiza PabelloNo ratings yet
- Molecular Identification Methods of Fish Species RDocument30 pagesMolecular Identification Methods of Fish Species RRhaiza PabelloNo ratings yet
- Lancet OncologyDocument8 pagesLancet OncologyRhaiza PabelloNo ratings yet
- Yusup 2020 IOP Conf. Ser. Earth Environ. Sci. 429 012046Document9 pagesYusup 2020 IOP Conf. Ser. Earth Environ. Sci. 429 012046Rhaiza PabelloNo ratings yet
- Search ResultsDocument28 pagesSearch ResultsRhaiza PabelloNo ratings yet
- IOP Conference Series: Earth and Environmental Science Identification and InventoryDocument6 pagesIOP Conference Series: Earth and Environmental Science Identification and InventoryRhaiza PabelloNo ratings yet
- DNA Barcoding As A Tool For Coral Reef ConservatioDocument14 pagesDNA Barcoding As A Tool For Coral Reef ConservatioRhaiza PabelloNo ratings yet
- Journal of Experimental and Clinical Cancer ResearchDocument23 pagesJournal of Experimental and Clinical Cancer ResearchRhaiza PabelloNo ratings yet
- Radiotherapy and OncologyDocument12 pagesRadiotherapy and OncologyRhaiza PabelloNo ratings yet
- 9 - Journal of CarcinogenesisDocument6 pages9 - Journal of CarcinogenesisRhaiza PabelloNo ratings yet
- 4 - Clinical Cancer ResearchDocument4 pages4 - Clinical Cancer ResearchRhaiza PabelloNo ratings yet
- 11 - Molecular OncologyDocument15 pages11 - Molecular OncologyRhaiza PabelloNo ratings yet
- 5 - Molecular CancerDocument6 pages5 - Molecular CancerRhaiza PabelloNo ratings yet
- Tfap2A Potentiates Lung Adenocarcinoma Metastasis By A Novel Mir-16 Family/Tfap2A/Psg9/ Tgf-Β Signaling PathwayDocument13 pagesTfap2A Potentiates Lung Adenocarcinoma Metastasis By A Novel Mir-16 Family/Tfap2A/Psg9/ Tgf-Β Signaling PathwayRhaiza PabelloNo ratings yet
- 3 - Nature Reviews Clinical OncologyDocument8 pages3 - Nature Reviews Clinical OncologyRhaiza PabelloNo ratings yet
- Exercises On DefinitionDocument1 pageExercises On DefinitionRhaiza PabelloNo ratings yet
- Current and Resistance ExerciseDocument2 pagesCurrent and Resistance ExerciseRhaiza PabelloNo ratings yet
- Physics 2 ECQ Quiz 1 - Current and ResistanceDocument2 pagesPhysics 2 ECQ Quiz 1 - Current and ResistanceRhaiza PabelloNo ratings yet
- Physics Trivia QuestionsDocument3 pagesPhysics Trivia QuestionsRhaiza PabelloNo ratings yet
- DAILY MOOD CHART: MONTH OF - : High +3 +2 +1Document2 pagesDAILY MOOD CHART: MONTH OF - : High +3 +2 +1Rhaiza PabelloNo ratings yet
- Pabello - Conceptual Questions On MagnetismDocument1 pagePabello - Conceptual Questions On MagnetismRhaiza PabelloNo ratings yet
- Pabello, Rhaiza P. April 28, 2020 Internet Exercise C1A1-YDocument3 pagesPabello, Rhaiza P. April 28, 2020 Internet Exercise C1A1-YRhaiza PabelloNo ratings yet
- The Role of Humanities and The Arts in 21st Century EducationDocument2 pagesThe Role of Humanities and The Arts in 21st Century EducationRhaiza PabelloNo ratings yet
- War of CurrentsDocument2 pagesWar of CurrentsRhaiza PabelloNo ratings yet
- DOST Scholarship FormDocument3 pagesDOST Scholarship FormRhaiza PabelloNo ratings yet
- Pabello - Conceptual Questions On MagnetismDocument1 pagePabello - Conceptual Questions On MagnetismRhaiza PabelloNo ratings yet
- FAR EASTERN UNIVERSITY ECQ Quiz 1Document3 pagesFAR EASTERN UNIVERSITY ECQ Quiz 1Rhaiza PabelloNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Anatomy, Physiology, and Pathophysiology of Muscular SystemDocument55 pagesThe Anatomy, Physiology, and Pathophysiology of Muscular SystemAnisaNo ratings yet
- LearnerDocument7 pagesLearnersudhacarhrNo ratings yet
- Ancient Healing Rods Reveal Secrets of PharaohsDocument28 pagesAncient Healing Rods Reveal Secrets of PharaohsMiguel Angelum100% (4)
- Book of Mythical BeastsDocument32 pagesBook of Mythical BeastsRishabhNo ratings yet
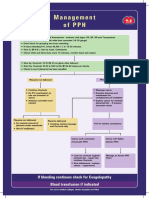
- Management of PPHDocument1 pageManagement of PPH098 U.KARTHIK SARAVANA KANTHNo ratings yet
- Analisis Kualitatif Dan Kuantitatif Kandungan Kimia Dari Ekstrak Heksan, Aseton, Etanol Dan Air Dari Umbi Bawang Putih (Allium Sativum Linn.)Document11 pagesAnalisis Kualitatif Dan Kuantitatif Kandungan Kimia Dari Ekstrak Heksan, Aseton, Etanol Dan Air Dari Umbi Bawang Putih (Allium Sativum Linn.)Tari PratiwiNo ratings yet
- E-Techno: Cbse Class-Ix - E6 - E-Techno JEE TEST DATE: 17-11-2020Document16 pagesE-Techno: Cbse Class-Ix - E6 - E-Techno JEE TEST DATE: 17-11-2020Himanshu ThakurNo ratings yet
- Morphology of Flowering PlantsDocument13 pagesMorphology of Flowering Plantschiragbhaskar16No ratings yet
- Gene TherapyDocument1 pageGene TherapyJenevieve B. CañeteNo ratings yet
- Warlock of The Magus World - Arc 2Document808 pagesWarlock of The Magus World - Arc 2Hitsuin MoviesNo ratings yet
- Nervous Responses of EarthwormDocument27 pagesNervous Responses of EarthwormKayla JoezetteNo ratings yet
- Gazzaniga The Ethical - Brain.the - Science.of - Our.moral - DilemmasDocument229 pagesGazzaniga The Ethical - Brain.the - Science.of - Our.moral - Dilemmaskid_latigo100% (9)
- Chapter 5Document14 pagesChapter 5Caitlin G.No ratings yet
- Ficha Técnica SpotcheckDocument5 pagesFicha Técnica SpotcheckJuan PazNo ratings yet
- Ap Environmental Science Free-Response QuestionsDocument6 pagesAp Environmental Science Free-Response Questionsapi-234419767No ratings yet
- RT-PCR Kit for RNA Detection up to 6.5kbDocument14 pagesRT-PCR Kit for RNA Detection up to 6.5kbLatifa Putri FajrNo ratings yet
- 0610 IECR P3 Q1 v2 PDFDocument9 pages0610 IECR P3 Q1 v2 PDFGAIA Educators - Home TuitionNo ratings yet
- Lipid Movement BiochemDocument9 pagesLipid Movement BiochemCrowNo ratings yet
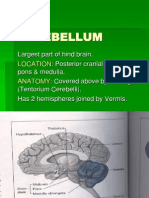
- Physiology of Cerebellum Lec Foe 2nd Year by DR Sadia Uploaded by ZaighamDocument61 pagesPhysiology of Cerebellum Lec Foe 2nd Year by DR Sadia Uploaded by ZaighamZaigham HammadNo ratings yet
- Paul Atriedes (PL 6/181pp) : Bene Gesserit Training (Container, Active 11)Document2 pagesPaul Atriedes (PL 6/181pp) : Bene Gesserit Training (Container, Active 11)dfgdfNo ratings yet
- Biology: Pearson EdexcelDocument24 pagesBiology: Pearson EdexcelAhmad MohdNo ratings yet
- Post Harvest Handling in LitchiDocument52 pagesPost Harvest Handling in LitchiDr Parag B JadhavNo ratings yet
- 03 Chapter 3Document148 pages03 Chapter 3arghaNo ratings yet
- Cosmeceutic ALS: Cosmetics and PharmaceuticalsDocument19 pagesCosmeceutic ALS: Cosmetics and PharmaceuticalssowjanyaNo ratings yet
- MOKUSAKU APPLICATIONS (Version 1)Document1 pageMOKUSAKU APPLICATIONS (Version 1)api-19621568No ratings yet
- Diencephalon PresentationDocument19 pagesDiencephalon Presentation300rNo ratings yet
- 1 s2.0 S0141022999001556 MainDocument21 pages1 s2.0 S0141022999001556 MainRenata ScoralickNo ratings yet
- Amazing Facts - Exercises 1Document3 pagesAmazing Facts - Exercises 1Alina DorobatNo ratings yet
- Colon 2014597 D FHTDocument4 pagesColon 2014597 D FHTyoohooNo ratings yet
- Week 6. Definision and DictionaryDocument19 pagesWeek 6. Definision and DictionaryafdreliautariazizahNo ratings yet