You might also like
- Review: Cells of Origin in CancerDocument9 pagesReview: Cells of Origin in CancerJohn TikasNo ratings yet
- Ordinal Regression Analysis - Fitting The Proportional Odds ModelDocument15 pagesOrdinal Regression Analysis - Fitting The Proportional Odds ModelJohn TikasNo ratings yet
- L11Document17 pagesL11John TikasNo ratings yet
- The Times Winning MovesDocument116 pagesThe Times Winning MovesamuronegaduNo ratings yet
- 64 Great Chess GamesDocument306 pages64 Great Chess GamesManijak999100% (6)
- Logit From PrincetonDocument15 pagesLogit From PrincetonJudy DomingoNo ratings yet
- (Ebook - PDF - Mathematics) - Abstract AlgebraDocument113 pages(Ebook - PDF - Mathematics) - Abstract Algebrambardak100% (2)
- Modern and Postmodern Challenges To Game TheoryDocument34 pagesModern and Postmodern Challenges To Game TheoryJohn TikasNo ratings yet
- Advances in Networks ElectrophysiologyDocument21 pagesAdvances in Networks ElectrophysiologyJohn TikasNo ratings yet
- Sartorius Muscle Metastases From Urothelial Cell CarcinomaDocument3 pagesSartorius Muscle Metastases From Urothelial Cell CarcinomaJohn TikasNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5783)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (72)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- APP Ch.3 OutlineDocument5 pagesAPP Ch.3 OutlineAlliePalmerNo ratings yet
- Cell Biology Product and Research Guide PDFDocument164 pagesCell Biology Product and Research Guide PDFBasma Al-KershNo ratings yet
- 1 - Brief Biography of Charles Darwin (1809 - 1882)Document22 pages1 - Brief Biography of Charles Darwin (1809 - 1882)vladNo ratings yet
- General Pathology (Lecture-1) - 3Document14 pagesGeneral Pathology (Lecture-1) - 3Usman AkramNo ratings yet
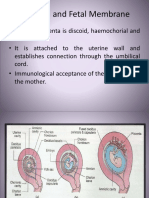
- Placenta and Fetal Membrane-6614Document45 pagesPlacenta and Fetal Membrane-6614Incredible DivineNo ratings yet
- Curly Hair Genetics - Dominant or RecessiveDocument4 pagesCurly Hair Genetics - Dominant or RecessiveimnylaroseNo ratings yet
- Aflp 1Document55 pagesAflp 1Purushotham PaspuletiNo ratings yet
- Growing Poultry Industry Meets Need for Alternative FeedsDocument47 pagesGrowing Poultry Industry Meets Need for Alternative FeedsJakin Aia TapanganNo ratings yet
- Cladogram CH 19 Part ADocument3 pagesCladogram CH 19 Part AJasmine lee100% (1)
- 1989 - Alterations of Life Span in The Nematode Caenorhabditis Elegans Under Monoxenic Culture ConditionsDocument14 pages1989 - Alterations of Life Span in The Nematode Caenorhabditis Elegans Under Monoxenic Culture ConditionsRaymond LaBoyNo ratings yet
- Pond EcosystemDocument13 pagesPond EcosystemTorres Wong Joeanne100% (1)
- Pest ManagementDocument5 pagesPest Managementkiran100% (1)
- Osmolarity&tonicitypracticeproblemsfrominternetDocument14 pagesOsmolarity&tonicitypracticeproblemsfrominternetbsktblbabe88No ratings yet
- Mader/Biology, 11/e - Chapter Outline: 3.1 Organic MoleculesDocument6 pagesMader/Biology, 11/e - Chapter Outline: 3.1 Organic Moleculesapi-455371000No ratings yet
- Food Poisoning - Outbreaks, Bacterial Sources and Adverse Health Effects (2015) PDFDocument294 pagesFood Poisoning - Outbreaks, Bacterial Sources and Adverse Health Effects (2015) PDFAndré Campelo100% (1)
- Hematology: Mohamad H Qari, MD, FRCPADocument49 pagesHematology: Mohamad H Qari, MD, FRCPASantoz ArieNo ratings yet
- OCR A A-Level Biology Retrieval Roulette COMPLETEDocument115 pagesOCR A A-Level Biology Retrieval Roulette COMPLETEtikif31811No ratings yet
- Effects of High Temperature On Nodulation and Nitrogen Fixation byDocument8 pagesEffects of High Temperature On Nodulation and Nitrogen Fixation byFábio Luís MostassoNo ratings yet
- 10 1126@science Aau3644Document14 pages10 1126@science Aau3644nicolasNo ratings yet
- Kementerian Agama Republik Indonesia: Penilaian Akhir Semester Ganjil Madrasah Aliyah TAHUN PELAJARAN 2020-2021Document7 pagesKementerian Agama Republik Indonesia: Penilaian Akhir Semester Ganjil Madrasah Aliyah TAHUN PELAJARAN 2020-2021Aisyah DNo ratings yet
- Histologi Usus Ikan NilaDocument14 pagesHistologi Usus Ikan NilaOnnaSafuraNo ratings yet
- Bioreaction Engineering Principles Nielsen-VilladsenDocument554 pagesBioreaction Engineering Principles Nielsen-VilladsenTonatiuh CortesNo ratings yet
- Tri-Fold TemplateDocument2 pagesTri-Fold Templateapi-260948905No ratings yet
- Ethics Ivf Pgc2004Document32 pagesEthics Ivf Pgc2004witch_26452No ratings yet
- Friedreich 'S Ataxia: Clinical Features, Pathogenesis and ManagementDocument12 pagesFriedreich 'S Ataxia: Clinical Features, Pathogenesis and ManagementVaibhav BhatiaNo ratings yet
- Chap 12 Energy and RespirationDocument34 pagesChap 12 Energy and RespirationGeorge ApidiNo ratings yet
- Biology Lesson 3Document5 pagesBiology Lesson 3Sean Mevrick VegaNo ratings yet
- NIH Public Access: Olfaction in Parkinson's Disease and Related DisordersDocument63 pagesNIH Public Access: Olfaction in Parkinson's Disease and Related DisordersCarmen CiursaşNo ratings yet
- Patentability of Biotech Subject MatterDocument136 pagesPatentability of Biotech Subject MatterGareth WealeNo ratings yet
- Bryophyta Water Relations PDFDocument40 pagesBryophyta Water Relations PDFCDB 1st Semester 2077No ratings yet