You might also like
- Yoshida 2016 Science Ideonella Sakaiensis Grows On PETDocument5 pagesYoshida 2016 Science Ideonella Sakaiensis Grows On PETAnonymous 0zrCNQNo ratings yet
- Characterization and Engineering of A Plastic-Degrading Aromatic PolyesteraseDocument8 pagesCharacterization and Engineering of A Plastic-Degrading Aromatic PolyesteraseUssyy NurhidayahNo ratings yet
- Unicaciones de La Naturaleza WordDocument8 pagesUnicaciones de La Naturaleza Wordhamilton gavilanezNo ratings yet
- Pet DegradaciónDocument4 pagesPet DegradaciónAntuanet ToledoNo ratings yet
- Polymers 13 02878 v2Document15 pagesPolymers 13 02878 v2DhaneshNo ratings yet
- Biodegradability of Plastics Challenges and MisconceptionsDocument3 pagesBiodegradability of Plastics Challenges and Misconceptionslucas112358No ratings yet
- Future Prospects: Role of Ideonella Sakaiensis in Reducing Plastic Waste: Current Updates andDocument6 pagesFuture Prospects: Role of Ideonella Sakaiensis in Reducing Plastic Waste: Current Updates andShadma KhanNo ratings yet
- ChemosphereDocument13 pagesChemosphereyunpu jiaNo ratings yet
- Environmental Clean-Up: Group 5-BiologyDocument8 pagesEnvironmental Clean-Up: Group 5-BiologyTrexi Mag-asoNo ratings yet
- Biodegradability of Plastics Challenges and MisconceptionsDocument3 pagesBiodegradability of Plastics Challenges and MisconceptionsShivam SamaiyaNo ratings yet
- Fmicb 12 751571Document13 pagesFmicb 12 751571Tron TronNo ratings yet
- Acs Est 7b04051Document3 pagesAcs Est 7b04051NIKHIL YEMULNo ratings yet
- 83-HelmholzPETRECYCLING Mueller 3 Enzyme PDFDocument5 pages83-HelmholzPETRECYCLING Mueller 3 Enzyme PDFLuis Guillermo Feria BarreraNo ratings yet
- SOAS Plast'Innov Promo2022Document6 pagesSOAS Plast'Innov Promo2022Kilian DuchesneNo ratings yet
- 10 1016@j Carbpol 2017 12 067Document53 pages10 1016@j Carbpol 2017 12 067Sabrine SellimiNo ratings yet
- Novel Plastic-Degrading EnzymesDocument10 pagesNovel Plastic-Degrading EnzymesPodria NationStatesNo ratings yet
- Brief Report Agar Plate-Based Screening Methods For The Identi Fication of Polyester Hydrolysis by Pseudomonas SpeciesDocument11 pagesBrief Report Agar Plate-Based Screening Methods For The Identi Fication of Polyester Hydrolysis by Pseudomonas SpeciesBodhi DharmaNo ratings yet
- Complete Bio-Degradation of Poly (Butylene Adipate-Co-Terephthalate) Via Engineered CutinasesDocument8 pagesComplete Bio-Degradation of Poly (Butylene Adipate-Co-Terephthalate) Via Engineered CutinasesShivani BehareNo ratings yet
- 1 s2.0 S030438942200471X MainDocument11 pages1 s2.0 S030438942200471X MainJulianna PeixotoNo ratings yet
- 1 s2.0 S0048969719359261 MainDocument8 pages1 s2.0 S0048969719359261 MainJULIANA ANDREA BETANCUR VELASQUEZNo ratings yet
- Paper Jan 2016Document7 pagesPaper Jan 2016Syed babar ali shahNo ratings yet
- Fbaccino,+phycol 2 1 62Document6 pagesFbaccino,+phycol 2 1 62Raksha MukhiaNo ratings yet
- New Perspectives in Plastic Biodegradation PDFDocument6 pagesNew Perspectives in Plastic Biodegradation PDFCristina RangelNo ratings yet
- Biocomposites of Alfa Fibers Dispersed in The Mater-Bi® Type BioplasticDocument35 pagesBiocomposites of Alfa Fibers Dispersed in The Mater-Bi® Type BioplastichanjunyieeNo ratings yet
- PETase Modification With Integration of MHETaseDocument6 pagesPETase Modification With Integration of MHETaseKilian DuchesneNo ratings yet
- Letters Applied Microbiology - 2020 - Wang - Enhancing Secretion of Polyethylene Terephthalate Hydrolase PETase in BacillusDocument7 pagesLetters Applied Microbiology - 2020 - Wang - Enhancing Secretion of Polyethylene Terephthalate Hydrolase PETase in Bacillusxuanzhengxie726No ratings yet
- Eduardo L. Almeida, Andrés Felipe Carrillo Rincón, Stephen A. Jackson and Alan D. W. DobsonDocument14 pagesEduardo L. Almeida, Andrés Felipe Carrillo Rincón, Stephen A. Jackson and Alan D. W. DobsonBodhi DharmaNo ratings yet
- Bioplastic From Pectin of Dragon Fruit HylocereusDocument6 pagesBioplastic From Pectin of Dragon Fruit HylocereusJulius EstacioNo ratings yet
- PLA Recent Overview 2010Document9 pagesPLA Recent Overview 2010庄妍张No ratings yet
- Biodegradation of Poly (Lactic Acid) - Starch-Coir Biocomposites Under Controlled Composting ConditionsDocument11 pagesBiodegradation of Poly (Lactic Acid) - Starch-Coir Biocomposites Under Controlled Composting ConditionsLong LeNo ratings yet
- Biorremediacion PETDocument15 pagesBiorremediacion PETezeNo ratings yet
- Optimization of Protoplast Isolation From The Gametophytes of Brown Alga Undaria Pinnatifida Using Response Surface MethodologyDocument13 pagesOptimization of Protoplast Isolation From The Gametophytes of Brown Alga Undaria Pinnatifida Using Response Surface MethodologyJo CynthiaNo ratings yet
- International Biodeterioration & Biodegradation: Miriam Santo, Ronen Weitsman, Alex SivanDocument7 pagesInternational Biodeterioration & Biodegradation: Miriam Santo, Ronen Weitsman, Alex SivanZero HeroNo ratings yet
- Extraction and Physicochemical Characterization of Chitin and Chitosan Isolated From House CricketDocument14 pagesExtraction and Physicochemical Characterization of Chitin and Chitosan Isolated From House CricketagnitaNo ratings yet
- Fieldtestonthebiodegradationofpolybutyleneadipate-Co-Terephthalatebasedmulchfilmsinsoil - Not Read YetDocument8 pagesFieldtestonthebiodegradationofpolybutyleneadipate-Co-Terephthalatebasedmulchfilmsinsoil - Not Read YetShailajhaNo ratings yet
- Biodegradation of Poly (Ethylene Terephthalate) by Bacillus SubtilisDocument4 pagesBiodegradation of Poly (Ethylene Terephthalate) by Bacillus SubtilisExon PerezNo ratings yet
- 1 s2.0 S1096717621000471 MainDocument13 pages1 s2.0 S1096717621000471 MainJorge Lima PérezNo ratings yet
- New Insights Into The Function and Global Distribution ofDocument34 pagesNew Insights Into The Function and Global Distribution ofWi FuentesNo ratings yet
- Screening and Characterization of Novel Lipase ProducingDocument24 pagesScreening and Characterization of Novel Lipase ProducingAqsa khanNo ratings yet
- Biomolecules 13 00441 v2Document23 pagesBiomolecules 13 00441 v2Sarah SalaNo ratings yet
- Carbonyll CompoundDocument13 pagesCarbonyll CompoundMafatih WebNo ratings yet
- Polymer Testing - 30 - 2010 - 60-68Document9 pagesPolymer Testing - 30 - 2010 - 60-68Carinna Saldaña - PierardNo ratings yet
- PET Polymer Recycling: AccessDocument3 pagesPET Polymer Recycling: AccessSanem Hazal TürkayNo ratings yet
- Ideonella Sakaiensis Sp. Nov., Isolated From A Microbial Consortium That Degrades Poly (Ethylene Terephthalate)Document6 pagesIdeonella Sakaiensis Sp. Nov., Isolated From A Microbial Consortium That Degrades Poly (Ethylene Terephthalate)SP GUPTANo ratings yet
- A Marine Bacterial Community Capable of Degrading Poly (Ethylene Terephthalate) and PolyethyleneDocument12 pagesA Marine Bacterial Community Capable of Degrading Poly (Ethylene Terephthalate) and PolyethyleneTron TronNo ratings yet
- 2010 - High-Density Polyethylene (HDPE) - Degrading PotentialDocument7 pages2010 - High-Density Polyethylene (HDPE) - Degrading PotentialHoa NắngNo ratings yet
- Extraction and Physico - IbitoyeDocument13 pagesExtraction and Physico - Ibitoyeruben alvaradoNo ratings yet
- Chemosphere: Navid Taghavi, Naresh Singhal, Wei-Qin Zhuang, Saeid BaroutianDocument14 pagesChemosphere: Navid Taghavi, Naresh Singhal, Wei-Qin Zhuang, Saeid BaroutianHoa NắngNo ratings yet
- New Advancements of Bioplastics in Medical ApplicationsDocument15 pagesNew Advancements of Bioplastics in Medical ApplicationsBrizeth García DíazNo ratings yet
- Extraction and Characterization of Coconut (Cocos Nucifera L.) Coir DustDocument9 pagesExtraction and Characterization of Coconut (Cocos Nucifera L.) Coir DustIgbinwole Kehinde SolomonNo ratings yet
- Degradation of Plastic by FungusDocument12 pagesDegradation of Plastic by Funguscindialim80No ratings yet
- 10 1002@app 50020Document13 pages10 1002@app 50020dhy182No ratings yet
- Carbonization of Animal Bone PDFDocument15 pagesCarbonization of Animal Bone PDFENDALKACHEW KERIENo ratings yet
- Synthesis and Characterization of Gelatin Based Polyester Urethane ScaffoldDocument10 pagesSynthesis and Characterization of Gelatin Based Polyester Urethane ScaffoldPaoloMMNo ratings yet
- Síntesis y Caracterización de Elastómeros de Poliuretano a Base de Quitosano y Poli (ε ‐ Caprolactona)Document9 pagesSíntesis y Caracterización de Elastómeros de Poliuretano a Base de Quitosano y Poli (ε ‐ Caprolactona)Neider Andres Vera SilvaNo ratings yet
- 1 s2.0 S2666893921001419 Main1Document9 pages1 s2.0 S2666893921001419 Main1jadeshades12No ratings yet
- 1 s2.0 S2238785421004877 MainDocument29 pages1 s2.0 S2238785421004877 MainLê Văn HuấnNo ratings yet
- Kerimat Complete Project 2Document39 pagesKerimat Complete Project 2TemidayoNo ratings yet
- Intensification of Yam-Starch Based Biodegradable Bioplastic Film With Bentonite For Food Packaging Application (2022)Document12 pagesIntensification of Yam-Starch Based Biodegradable Bioplastic Film With Bentonite For Food Packaging Application (2022)Mahmoud BelabbassiNo ratings yet
- Molecular Breeding and Genetics of Applied MicroorganismsFrom EverandMolecular Breeding and Genetics of Applied MicroorganismsKenji SakaguchiNo ratings yet
- Solubility Product ConstantsDocument6 pagesSolubility Product ConstantsBilal AhmedNo ratings yet
- Midi Pro Adapter ManualDocument34 pagesMidi Pro Adapter ManualUli ZukowskiNo ratings yet
- BARUDocument53 pagesBARUhueuaNo ratings yet
- Hazardous Locations: C.E.C. ClassificationsDocument4 pagesHazardous Locations: C.E.C. ClassificationsThananuwat SuksaroNo ratings yet
- Smart Protein Plant Based Food Sector Report 2Document199 pagesSmart Protein Plant Based Food Sector Report 2campeon00magnatesNo ratings yet
- Contoh Discussion Text Tentang HomeworkDocument8 pagesContoh Discussion Text Tentang Homeworkg3p35rs6100% (1)
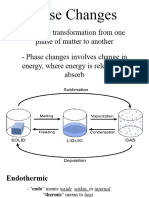
- General Chemistry 2 Q1 Lesson 5 Endothermic and Exotheric Reaction and Heating and Cooling CurveDocument19 pagesGeneral Chemistry 2 Q1 Lesson 5 Endothermic and Exotheric Reaction and Heating and Cooling CurveJolo Allexice R. PinedaNo ratings yet
- Low Budget Music Promotion and PublicityDocument41 pagesLow Budget Music Promotion and PublicityFola Folayan100% (3)
- Power Curbers, Inc. v. E. D. Etnyre & Co. and A. E. Finley & Associates, Inc., 298 F.2d 484, 4th Cir. (1962)Document18 pagesPower Curbers, Inc. v. E. D. Etnyre & Co. and A. E. Finley & Associates, Inc., 298 F.2d 484, 4th Cir. (1962)Scribd Government DocsNo ratings yet
- Chapter 4 - Basic ProbabilityDocument37 pagesChapter 4 - Basic Probabilitynadya shafirahNo ratings yet
- RMC 102-2017 HighlightsDocument3 pagesRMC 102-2017 HighlightsmmeeeowwNo ratings yet
- Aditya Birla GroupDocument21 pagesAditya Birla GroupNarendra ThummarNo ratings yet
- Comparative Study Between Online and Offilne Learning With Reference of Tutedude E-LearningDocument61 pagesComparative Study Between Online and Offilne Learning With Reference of Tutedude E-LearningDeeksha Saxena0% (2)
- Cyclic MeditationDocument8 pagesCyclic MeditationSatadal GuptaNo ratings yet
- Seabank Statement 20220726Document4 pagesSeabank Statement 20220726Alesa WahabappNo ratings yet
- Jan 25th 6 TicketsDocument2 pagesJan 25th 6 TicketsMohan Raj VeerasamiNo ratings yet
- Odisha State Museum-1Document26 pagesOdisha State Museum-1ajitkpatnaikNo ratings yet
- Stentofon Pulse: IP Based Intercom SystemDocument22 pagesStentofon Pulse: IP Based Intercom SystemCraigNo ratings yet
- Artificial Intelligence Techniques For Encrypt Images Based On The Chaotic System Implemented On Field-Programmable Gate ArrayDocument10 pagesArtificial Intelligence Techniques For Encrypt Images Based On The Chaotic System Implemented On Field-Programmable Gate ArrayIAES IJAINo ratings yet
- Soujanya Reddy (New)Document6 pagesSoujanya Reddy (New)durgaNo ratings yet
- Introduction To Atomistic Simulation Through Density Functional TheoryDocument21 pagesIntroduction To Atomistic Simulation Through Density Functional TheoryTarang AgrawalNo ratings yet
- Practical Modern SCADA Protocols. DNP3, 60870.5 and Related SystemsDocument4 pagesPractical Modern SCADA Protocols. DNP3, 60870.5 and Related Systemsalejogomez200No ratings yet
- Mozal Finance EXCEL Group 15dec2013Document15 pagesMozal Finance EXCEL Group 15dec2013Abhijit TailangNo ratings yet
- Satish Gujral - FinalDocument23 pagesSatish Gujral - Finalsatya madhuNo ratings yet
- in 30 MinutesDocument5 pagesin 30 MinutesCésar DiazNo ratings yet
- Application of Geoelectric Method For GroundwaterDocument11 pagesApplication of Geoelectric Method For GroundwaterMunther DhahirNo ratings yet
- Chapter 2Document8 pagesChapter 2Fahmia MidtimbangNo ratings yet
- 1916 South American Championship Squads - WikipediaDocument6 pages1916 South American Championship Squads - WikipediaCristian VillamayorNo ratings yet
- MME 52106 - Optimization in Matlab - NN ToolboxDocument14 pagesMME 52106 - Optimization in Matlab - NN ToolboxAdarshNo ratings yet
- TFGDocument46 pagesTFGAlex Gigena50% (2)