You might also like
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Antibiotics Resistence PDFDocument45 pagesAntibiotics Resistence PDFOsiithaa CañaszNo ratings yet
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Infectous Endocarditis PDFDocument22 pagesInfectous Endocarditis PDFOsiithaa CañaszNo ratings yet
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- Vitiliggo PDFDocument11 pagesVitiliggo PDFOsiithaa CañaszNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Impetigo, Erysipelas PDFDocument18 pagesImpetigo, Erysipelas PDFOsiithaa CañaszNo ratings yet
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Pulmonary Hypertension All PeopleDocument19 pagesPulmonary Hypertension All PeopleOsiithaa CañaszNo ratings yet
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- SX KlinefelterDocument14 pagesSX KlinefelterOsiithaa CañaszNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Outpatient Emergencies Anaphylaxis PDFDocument16 pagesOutpatient Emergencies Anaphylaxis PDFOsiithaa CañaszNo ratings yet
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Aih 4 PDFDocument14 pagesAih 4 PDFOsiithaa CañaszNo ratings yet
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Acute Lymphoblastic Leukemia Review 2017 PDFDocument12 pagesAcute Lymphoblastic Leukemia Review 2017 PDFOsiithaa CañaszNo ratings yet
- Cardiac Emergencies in Neonates and Young InfantsDocument6 pagesCardiac Emergencies in Neonates and Young InfantsOsiithaa CañaszNo ratings yet
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- 5Document2 pages5Osiithaa CañaszNo ratings yet
- Ijerph 13 01003 PDFDocument10 pagesIjerph 13 01003 PDFOsiithaa CañaszNo ratings yet
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Case Report: A Comprehensive Neuropsychological Assessment of A Case of Super Ficial SiderosisDocument8 pagesCase Report: A Comprehensive Neuropsychological Assessment of A Case of Super Ficial SiderosisOsiithaa CañaszNo ratings yet
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Disseminated Intravascular Coagulation PDFDocument16 pagesDisseminated Intravascular Coagulation PDFOsiithaa CañaszNo ratings yet
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- Basic Shock Physiology PDFDocument14 pagesBasic Shock Physiology PDFOsiithaa CañaszNo ratings yet
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- An Immunological Perspective On Neonatal Sepsis PDFDocument22 pagesAn Immunological Perspective On Neonatal Sepsis PDFOsiithaa CañaszNo ratings yet
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- CholedocholithiasisDocument9 pagesCholedocholithiasisOsiithaa CañaszNo ratings yet
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- Infectous Endocarditis PDFDocument22 pagesInfectous Endocarditis PDFOsiithaa CañaszNo ratings yet
- The Metabolic Syndrome and Chronic KidneyDocument11 pagesThe Metabolic Syndrome and Chronic KidneyOsiithaa CañaszNo ratings yet
- Pulmonary Hypertension Due To Left Heart PDFDocument12 pagesPulmonary Hypertension Due To Left Heart PDFOsiithaa CañaszNo ratings yet
- The Metabolic Syndrome and Chronic KidneyDocument11 pagesThe Metabolic Syndrome and Chronic KidneyOsiithaa CañaszNo ratings yet
- Reportinverbs PDFDocument3 pagesReportinverbs PDFOsiithaa CañaszNo ratings yet
- An Immunological Perspective On Neonatal Sepsis PDFDocument22 pagesAn Immunological Perspective On Neonatal Sepsis PDFOsiithaa CañaszNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Review of New Hypertension Guidelines PDFDocument4 pagesReview of New Hypertension Guidelines PDFOsiithaa CañaszNo ratings yet
- Pathophysiology Management of Alcoholic Liver 2016Document16 pagesPathophysiology Management of Alcoholic Liver 2016Osiithaa CañaszNo ratings yet
- JNC 8Document14 pagesJNC 8amiwahyuniNo ratings yet
- Pediatric SepsisDocument16 pagesPediatric SepsisOsiithaa CañaszNo ratings yet
- Identification and Management of Alcohol Withdrawal SyndromeDocument13 pagesIdentification and Management of Alcohol Withdrawal SyndromeOsiithaa CañaszNo ratings yet
- Neonatal SepsisDocument8 pagesNeonatal SepsisOsiithaa CañaszNo ratings yet
- Hypertensive Emergency PDFDocument14 pagesHypertensive Emergency PDFOsiithaa CañaszNo ratings yet
- Detecting Cancer Spread Through Peritoneal Fluid AnalysisDocument16 pagesDetecting Cancer Spread Through Peritoneal Fluid AnalysisnanxtoyahNo ratings yet
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Clotting Factors List and Mnemonics For Easy Learning - All Medical StuffDocument10 pagesClotting Factors List and Mnemonics For Easy Learning - All Medical Stuffmonika ramosNo ratings yet
- What You Always Wanted To Know About SclerologyDocument20 pagesWhat You Always Wanted To Know About Sclerologyscientist786No ratings yet
- Physical and Physiological Changes in PregnancyDocument35 pagesPhysical and Physiological Changes in PregnancyAYO NELSONNo ratings yet
- HumanPhysiology Oogenesis Spermatogenesis FertilizationDocument19 pagesHumanPhysiology Oogenesis Spermatogenesis FertilizationAlam SalamNo ratings yet
- BMEN 301 F2021 Module 3-The Mechanics of Soft TissueDocument18 pagesBMEN 301 F2021 Module 3-The Mechanics of Soft TissueAyesha ANo ratings yet
- Fundamentals of Anatomy and Physiology: Course Description and ObjectivesDocument3 pagesFundamentals of Anatomy and Physiology: Course Description and Objectivesvamsi karna100% (1)
- RBC and Bleeding Disorders Outline Explains Anemia, Hemolytic DisordersDocument19 pagesRBC and Bleeding Disorders Outline Explains Anemia, Hemolytic DisordersNatan BabekNo ratings yet
- The Integumentary System: Structure and Function of SkinDocument10 pagesThe Integumentary System: Structure and Function of SkinDonto KonoNo ratings yet
- Case Study of UTI - Urinary Tract InfectionDocument9 pagesCase Study of UTI - Urinary Tract InfectionArvin Ian Penaflor75% (8)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- MCU 2019 Essentials in Gynecology For Undergraduate Medical StudentsDocument414 pagesMCU 2019 Essentials in Gynecology For Undergraduate Medical StudentsHAVIZ YUAD100% (1)
- Pelvis and Perineum - Anatomy, Vessels, Nerves - KenhubDocument11 pagesPelvis and Perineum - Anatomy, Vessels, Nerves - KenhubDhika100% (1)
- Cell Structure and FunctionDocument10 pagesCell Structure and FunctionKanaga ValliNo ratings yet
- NUR11O1 Integrated Human Anatomy and Physiology Department of Biology Institute of Arts and Sciences Far Eastern UniversityDocument5 pagesNUR11O1 Integrated Human Anatomy and Physiology Department of Biology Institute of Arts and Sciences Far Eastern UniversityYda Maxine PalmaNo ratings yet
- Anatomy and Physiology of the Intrinsic Cardiac Autonomic Nervous SystemDocument5 pagesAnatomy and Physiology of the Intrinsic Cardiac Autonomic Nervous SystemSori B. Requena C.No ratings yet
- Science Grade 3 Life Unit 3L1Document22 pagesScience Grade 3 Life Unit 3L1Cristina Isabel Saavedra CuellesNo ratings yet
- Cellular Membranes: Structures and FunctionsDocument7 pagesCellular Membranes: Structures and FunctionsSwasti Agnihotri100% (1)
- JC FMH Revisited 1 The Role of Mechano - TransductionDocument31 pagesJC FMH Revisited 1 The Role of Mechano - Transductionchirag sahgalNo ratings yet
- Photosynthesis Diagrams WorksheetDocument2 pagesPhotosynthesis Diagrams WorksheetG Meera Prem Kumar20% (5)
- Microflora PericoronariteDocument45 pagesMicroflora PericoronariteCeresău MarinaNo ratings yet
- Amniotic Fluid PDFDocument24 pagesAmniotic Fluid PDFJoshua Ty CayetanoNo ratings yet
- Pocket Tutor ECG Interpretation PDFDocument165 pagesPocket Tutor ECG Interpretation PDFasri100% (3)
- The Structure and Functions of the NucleusDocument19 pagesThe Structure and Functions of the NucleusYuu Ayu'k LifestarNo ratings yet
- Antigen Antibody Reactions Part2 0 0Document26 pagesAntigen Antibody Reactions Part2 0 0Marl EstradaNo ratings yet
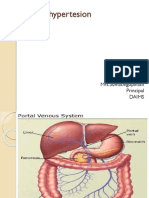
- Portal HypertensionDocument23 pagesPortal HypertensionSumathi GopinathNo ratings yet
- Percutaneous Interventional Cardiovascular Medicine - The PCR-EAPCI TextbookDocument29 pagesPercutaneous Interventional Cardiovascular Medicine - The PCR-EAPCI TextbookRanda Tabbah100% (1)
- KayleeDocument3 pagesKayleeKaylee RamsaroopNo ratings yet
- Surgery As An Aid To More Efficient Service With Prosthetic Dentures KazanjianDocument16 pagesSurgery As An Aid To More Efficient Service With Prosthetic Dentures KazanjianJuan Sierra ZambranoNo ratings yet
- Aubf TablesDocument24 pagesAubf TablesCanne OpadaNo ratings yet
- The Respiratory SystemDocument6 pagesThe Respiratory SystemIftikharNo ratings yet
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionFrom EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionRating: 4 out of 5 stars4/5 (402)
- Roxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingFrom EverandRoxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingRating: 5 out of 5 stars5/5 (4)
- The Age of Magical Overthinking: Notes on Modern IrrationalityFrom EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityRating: 4 out of 5 stars4/5 (13)
- The Happiness Trap: How to Stop Struggling and Start LivingFrom EverandThe Happiness Trap: How to Stop Struggling and Start LivingRating: 4 out of 5 stars4/5 (1)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeFrom EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeNo ratings yet
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsFrom EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsRating: 3.5 out of 5 stars3.5/5 (3)