You might also like
- Business Management System: Orrective Ction EportDocument2 pagesBusiness Management System: Orrective Ction EportBự BụngNo ratings yet
- Waldorf Curriculum ChartDocument1 pageWaldorf Curriculum Chartplanetalingua2020100% (1)
- Test Bank For Cognitive Psychology Connecting Mind Research and Everyday Experience 3rd Edition e Bruce GoldsteinDocument24 pagesTest Bank For Cognitive Psychology Connecting Mind Research and Everyday Experience 3rd Edition e Bruce GoldsteinMichaelThomasyqdi100% (49)
- Micropro CannDocument4 pagesMicropro CannJorge Carnicer OrujNo ratings yet
- Honda Ss50e General Export Crankcase - Schematic PartsficheDocument16 pagesHonda Ss50e General Export Crankcase - Schematic PartsficheBự BụngNo ratings yet
- Workshop Generic Hand Out Jan 2010Document15 pagesWorkshop Generic Hand Out Jan 2010Bự BụngNo ratings yet
- Cactus Tissue CultureDocument8 pagesCactus Tissue CultureHüseyin Avni TokgözNo ratings yet
- Cooking Oils and Smoke Points - What To Know and How To Choose The Right Cooking Oil - 2020 - MasterClassDocument7 pagesCooking Oils and Smoke Points - What To Know and How To Choose The Right Cooking Oil - 2020 - MasterClasschumbefredNo ratings yet
- Callo GenesisDocument8 pagesCallo GenesisJoselin Gisela Franco TapiaNo ratings yet
- tmpAE4A TMPDocument12 pagestmpAE4A TMPFrontiersNo ratings yet
- Papaya in VitroDocument5 pagesPapaya in VitroBiomontec Biomontec BiomontecNo ratings yet
- The in Uence of Phytohormones, Wheat Ovary Co-Culture, and Temperature Stress On Anther Culture Response of Watermelon (Citrullus Lanatus L.)Document11 pagesThe in Uence of Phytohormones, Wheat Ovary Co-Culture, and Temperature Stress On Anther Culture Response of Watermelon (Citrullus Lanatus L.)Hong Yee YongNo ratings yet
- Full TextDocument3 pagesFull TextarushwaniNo ratings yet
- Factors Affecting Haploid Induction Through in Vitro Gynogenesis in Summer Squash (Cucurbita Pepo L.)Document6 pagesFactors Affecting Haploid Induction Through in Vitro Gynogenesis in Summer Squash (Cucurbita Pepo L.)38 Sourav BhowmikNo ratings yet
- Ajol File Journals - 82 - Articles - 127451 - Submission - Proof - 127451 973 346045 1 10 20151214Document13 pagesAjol File Journals - 82 - Articles - 127451 - Submission - Proof - 127451 973 346045 1 10 20151214SilvanaNo ratings yet
- Tmp966e TMPDocument8 pagesTmp966e TMPFrontiersNo ratings yet
- Seed Dormancy in Red Rice' Plant Physiol. 1992 Footitt 1196 202Document7 pagesSeed Dormancy in Red Rice' Plant Physiol. 1992 Footitt 1196 202Phan Thúc ĐịnhNo ratings yet
- MCN 163Document11 pagesMCN 163Bự BụngNo ratings yet
- 140 MichaelDocument8 pages140 MichaelfatmawatiNo ratings yet
- C Elegans Krishnareddy - and - PullaiahDocument7 pagesC Elegans Krishnareddy - and - PullaiahDr. P V Krishna ReddyNo ratings yet
- Ijasr - Agri - In-Vitro Micropropagation of A Medicinal Plant Aparajita - Clitoria Ternatea L.Document6 pagesIjasr - Agri - In-Vitro Micropropagation of A Medicinal Plant Aparajita - Clitoria Ternatea L.TJPRC PublicationsNo ratings yet
- 8878-Article Text-31292-1-10-20131010Document6 pages8878-Article Text-31292-1-10-20131010ÖMER FARUK BORANo ratings yet
- An Efficient Micropropagation Protocol For Eclipta AlbaDocument6 pagesAn Efficient Micropropagation Protocol For Eclipta AlbaSuprabuddha KunduNo ratings yet
- KuljarDocument8 pagesKuljarRina Trihandayani PutriNo ratings yet
- Doubled Haploids in RiceDocument5 pagesDoubled Haploids in RiceZulfikar FauziNo ratings yet
- Protocol Optimisation For Micropropagation of Ethiopian YAMDocument15 pagesProtocol Optimisation For Micropropagation of Ethiopian YAMbahiran alwaysNo ratings yet
- A:1025084203958Document5 pagesA:1025084203958Rahul RanaNo ratings yet
- Ovule Culture To Obtain Progeny From Hybrid Seedless Bunch GrapesDocument6 pagesOvule Culture To Obtain Progeny From Hybrid Seedless Bunch GrapesDemar PeartNo ratings yet
- In Vitro Plant Regeneration From Narbon Vetch (Vicia Narbonensis L.) Using Cotyledonary Node ExplantsDocument4 pagesIn Vitro Plant Regeneration From Narbon Vetch (Vicia Narbonensis L.) Using Cotyledonary Node ExplantsAriana ChimiNo ratings yet
- Asymbiotic Seed Germination and in Vitro Seedling Development of Cyrtopodium Punctatum - A Propagation Protocol For An Endangered Florida Native OrchidDocument9 pagesAsymbiotic Seed Germination and in Vitro Seedling Development of Cyrtopodium Punctatum - A Propagation Protocol For An Endangered Florida Native OrchidOMAR RENTERIA MARTINNo ratings yet
- 10.1007/s11627 000 0052 0Document6 pages10.1007/s11627 000 0052 0Ana ResendeNo ratings yet
- Vinter Halter 2008Document7 pagesVinter Halter 2008María Reynel TarazonaNo ratings yet
- Passiflora in VitroDocument6 pagesPassiflora in VitroOskar GarciaNo ratings yet
- Micro-And Cutting Propagation of Silver Maple. I. Results With Adult and Juvenile PropagulesDocument7 pagesMicro-And Cutting Propagation of Silver Maple. I. Results With Adult and Juvenile Propagulesgazapo1221No ratings yet
- Red Sanders in Vitro Micropropagation MaduraiDocument6 pagesRed Sanders in Vitro Micropropagation MaduraiAPCCF RU BLORENo ratings yet
- A Study On Seed Germination and Seedling Development Of: Spathoglottis Bl. OrchidsDocument16 pagesA Study On Seed Germination and Seedling Development Of: Spathoglottis Bl. OrchidsBự BụngNo ratings yet
- A Wounding Method and Liquid CultureDocument6 pagesA Wounding Method and Liquid CulturehaibiotechNo ratings yet
- Micropropagation of Date Palm Phoenix DactyliferaDocument13 pagesMicropropagation of Date Palm Phoenix DactyliferaSelvakapoorNo ratings yet
- Thidiazuron Induced Somatic Embryogenesis and Plant Regeneration in Capsicum AnnuumDocument4 pagesThidiazuron Induced Somatic Embryogenesis and Plant Regeneration in Capsicum AnnuumAriana ChimiNo ratings yet
- Microspore-Derived Embryos From Quercus Suber Anthers Mimic Zygotic Embryos and Maintain Haploidy in Long-Term Anther CultureDocument8 pagesMicrospore-Derived Embryos From Quercus Suber Anthers Mimic Zygotic Embryos and Maintain Haploidy in Long-Term Anther CultureKristem KertzeifNo ratings yet
- Micropropagation and Germplasm Conservation of Four Chickpea (Cicer Arietinum L.) GenotypesDocument9 pagesMicropropagation and Germplasm Conservation of Four Chickpea (Cicer Arietinum L.) Genotypesbahiran alwaysNo ratings yet
- 2012 Chandra JPPRDocument6 pages2012 Chandra JPPRknchandu1No ratings yet
- Aahigh-Frequency Shoot Regeneration Throug PDFDocument9 pagesAahigh-Frequency Shoot Regeneration Throug PDFjammesNo ratings yet
- Effects of Strat in Polyethylene Glycol Soul On Germ of 3 N. Amercian Shrub SpeciesDocument10 pagesEffects of Strat in Polyethylene Glycol Soul On Germ of 3 N. Amercian Shrub SpeciesNassimaNo ratings yet
- Acido SulfuricoDocument8 pagesAcido Sulfuricoplanhigion06No ratings yet
- Adonis Vernalis 2Document4 pagesAdonis Vernalis 2amaliaNo ratings yet
- Somatic Embryogenesis and Enhanced Shoot Organogenesis In: Metabriggsia Ovalifolia W. T. WangDocument9 pagesSomatic Embryogenesis and Enhanced Shoot Organogenesis In: Metabriggsia Ovalifolia W. T. WangTica DuyênNo ratings yet
- MS 13-2009 e PrintDocument15 pagesMS 13-2009 e PrinteditorofijtosNo ratings yet
- Methods of Breaking Seed Dormancy For Ornamental Passion Fruit SpeciesDocument7 pagesMethods of Breaking Seed Dormancy For Ornamental Passion Fruit SpeciesNico RogofskiNo ratings yet
- tmpF930 TMPDocument9 pagestmpF930 TMPFrontiersNo ratings yet
- Different Treatment of Rice Seed Dormancy Breaking, Germination of Both Wild Species and Cultivated Varieties (Oryza Sativa L.)Document8 pagesDifferent Treatment of Rice Seed Dormancy Breaking, Germination of Both Wild Species and Cultivated Varieties (Oryza Sativa L.)Smr OnlyNo ratings yet
- Micropropagation of Eclipta Alba (Linn.) Hassk-A Valuable Medicinal HerbDocument8 pagesMicropropagation of Eclipta Alba (Linn.) Hassk-A Valuable Medicinal HerbprasathkrpNo ratings yet
- Somatic EmbryogenesisDocument8 pagesSomatic EmbryogenesisCarolina Torres GaleanoNo ratings yet
- Effect of Plant Growth Regulator On Micropropagtion of Ginger (Zingiber Officinale Rosc.) Cv-Suprava and SuruchiDocument15 pagesEffect of Plant Growth Regulator On Micropropagtion of Ginger (Zingiber Officinale Rosc.) Cv-Suprava and SuruchiMalek_h88No ratings yet
- 43 PDFDocument6 pages43 PDFkahkashan kazmiNo ratings yet
- Faba L.: Establishment of The Regeneration System For ViciaDocument8 pagesFaba L.: Establishment of The Regeneration System For ViciaAriana ChimiNo ratings yet
- In Vitro Propagation of Anisochilus Carnosus (LF.) Wallich - A Medicinal PlantDocument3 pagesIn Vitro Propagation of Anisochilus Carnosus (LF.) Wallich - A Medicinal PlantiaetsdiaetsdNo ratings yet
- Plant Regeneration From Petiole and Leaf of African Violet (Saintpaulia Ionantha Wendl.) Cultured in VitroDocument8 pagesPlant Regeneration From Petiole and Leaf of African Violet (Saintpaulia Ionantha Wendl.) Cultured in VitroYanely CanalesNo ratings yet
- Germination Behavior of Jatropha Curcas L. After Different Imbibition TimesDocument7 pagesGermination Behavior of Jatropha Curcas L. After Different Imbibition Timesrocio quispe cruzNo ratings yet
- Germination Behavior of Jatropha Curcas L. After Different Imbibition TimesDocument7 pagesGermination Behavior of Jatropha Curcas L. After Different Imbibition Timesp3dr0javaNo ratings yet
- Improvement of Asymbiotic Seed Germination and Seedling Development of Cypripedium Macranthos Sw. With Organic AdditivesDocument8 pagesImprovement of Asymbiotic Seed Germination and Seedling Development of Cypripedium Macranthos Sw. With Organic AdditivesBự BụngNo ratings yet
- EJDR - Volume 72 - Issue 1 - Pages 73-87Document15 pagesEJDR - Volume 72 - Issue 1 - Pages 73-87Magali Ruiz RivasNo ratings yet
- Rachmawati Et Al. 2020. Adventitious Shoots Derived From Leaf Explants in in Vitro Mass Propagation of Indonesian Selected Anthurium Clones (#674085) - 1145272Document8 pagesRachmawati Et Al. 2020. Adventitious Shoots Derived From Leaf Explants in in Vitro Mass Propagation of Indonesian Selected Anthurium Clones (#674085) - 1145272Budi WinartoNo ratings yet
- Karam 2000Document11 pagesKaram 2000Sheyla Zubiaga SaavedraNo ratings yet
- Via Suspension Cultures of Banana (Musa SPP.) : Highly Efficient Somatic Embryogenesis and Plant RegenerationDocument12 pagesVia Suspension Cultures of Banana (Musa SPP.) : Highly Efficient Somatic Embryogenesis and Plant Regenerationth_kiranNo ratings yet
- Acorns and Bitter Roots: Starch Grain Research in the Prehistoric Eastern WoodlandsFrom EverandAcorns and Bitter Roots: Starch Grain Research in the Prehistoric Eastern WoodlandsNo ratings yet
- Các Nguyên Tố Dinh Dưỡng Và Phân Bón Vi LượngDocument2 pagesCác Nguyên Tố Dinh Dưỡng Và Phân Bón Vi LượngBự BụngNo ratings yet
- Needed UL Docs - LABDocument20 pagesNeeded UL Docs - LABBự BụngNo ratings yet
- Improvement of Asymbiotic Seed Germination and Seedling Development of Cypripedium Macranthos Sw. With Organic AdditivesDocument8 pagesImprovement of Asymbiotic Seed Germination and Seedling Development of Cypripedium Macranthos Sw. With Organic AdditivesBự BụngNo ratings yet
- Article WJPR 1472956876Document14 pagesArticle WJPR 1472956876Bự BụngNo ratings yet
- MCN 163Document11 pagesMCN 163Bự BụngNo ratings yet
- Introduction To Plant Tissue Culture: Preliminary Workshop ProgramDocument7 pagesIntroduction To Plant Tissue Culture: Preliminary Workshop ProgramBự BụngNo ratings yet
- Instructions For Use: Hokkaido University Collection of Scholarly and Academic Papers: HUSCAPDocument14 pagesInstructions For Use: Hokkaido University Collection of Scholarly and Academic Papers: HUSCAPBự BụngNo ratings yet
- A Study On Seed Germination and Seedling Development Of: Spathoglottis Bl. OrchidsDocument16 pagesA Study On Seed Germination and Seedling Development Of: Spathoglottis Bl. OrchidsBự BụngNo ratings yet
- Comparison of Different Culture Media For The in Vitro Culture of Dendrobium (Orchidaceae)Document5 pagesComparison of Different Culture Media For The in Vitro Culture of Dendrobium (Orchidaceae)Bự BụngNo ratings yet
- 15 Vijayakumar Et AlDocument5 pages15 Vijayakumar Et AlBự BụngNo ratings yet
- Mass Propagation of Orchids Through in Vitro Seed Culture TechnologyDocument4 pagesMass Propagation of Orchids Through in Vitro Seed Culture TechnologyBự BụngNo ratings yet
- Mass Propagation of Orchids Through in Vitro Seed Culture TechnologyDocument4 pagesMass Propagation of Orchids Through in Vitro Seed Culture TechnologyBự BụngNo ratings yet
- Asymbiotic in Vitro Seed Germination and Regeneration of Vanda Coerulea Giff. Ex. Lindl., An Endangered Orchid From Northeast IndiaDocument5 pagesAsymbiotic in Vitro Seed Germination and Regeneration of Vanda Coerulea Giff. Ex. Lindl., An Endangered Orchid From Northeast IndiaBự BụngNo ratings yet
- Catalogue Cymbidium PDFDocument21 pagesCatalogue Cymbidium PDFBự BụngNo ratings yet
- Factors Affecting Seed Germination Of: Grammatophylum Speciosum Cultured in VitroDocument5 pagesFactors Affecting Seed Germination Of: Grammatophylum Speciosum Cultured in VitroBự BụngNo ratings yet
- Ho Chi Minh Hotel List 2010-04Document8 pagesHo Chi Minh Hotel List 2010-04Bự BụngNo ratings yet
- 8D Report: Issue/concern???: D4: Finding The Root Cause (Problem Analysis)Document2 pages8D Report: Issue/concern???: D4: Finding The Root Cause (Problem Analysis)Bự BụngNo ratings yet
- Potential Over-Etching Simulation ResultDocument5 pagesPotential Over-Etching Simulation ResultBự BụngNo ratings yet
- Test Report Submission WD ProgramsDocument1 pageTest Report Submission WD ProgramsBự BụngNo ratings yet
- Pyralux LF AdhesiveDocument2 pagesPyralux LF AdhesiveBự BụngNo ratings yet
- ViTrox 20230728 HLIBDocument4 pagesViTrox 20230728 HLIBkim heeNo ratings yet
- HitchjikersGuide v1Document126 pagesHitchjikersGuide v1ArushiNo ratings yet
- TG - Health 3 - Q3Document29 pagesTG - Health 3 - Q3LouieNo ratings yet
- Task 2 - The Nature of Linguistics and LanguageDocument8 pagesTask 2 - The Nature of Linguistics and LanguageValentina Cardenas VilleroNo ratings yet
- HSE Induction Training 1687407986Document59 pagesHSE Induction Training 1687407986vishnuvarthanNo ratings yet
- 12-Zoomlion 70t Crawler Crane Specs - v2.4Document2 pages12-Zoomlion 70t Crawler Crane Specs - v2.4Athul BabuNo ratings yet
- Sorsogon State College: Republic of The Philippines Bulan Campus Bulan, SorsogonDocument4 pagesSorsogon State College: Republic of The Philippines Bulan Campus Bulan, Sorsogonerickson hernanNo ratings yet
- National Healthy Lifestyle ProgramDocument6 pagesNational Healthy Lifestyle Programmale nurseNo ratings yet
- January Payslip 2023.pdf - 1-2Document1 pageJanuary Payslip 2023.pdf - 1-2Arbaz KhanNo ratings yet
- Tateni Home Care ServicesDocument2 pagesTateni Home Care ServicesAlejandro CardonaNo ratings yet
- Surefire Hellfighter Power Cord QuestionDocument3 pagesSurefire Hellfighter Power Cord QuestionPedro VianaNo ratings yet
- Types of Vegetation in Western EuropeDocument12 pagesTypes of Vegetation in Western EuropeChemutai EzekielNo ratings yet
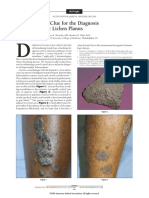
- A Novel Visual Clue For The Diagnosis of Hypertrophic Lichen PlanusDocument1 pageA Novel Visual Clue For The Diagnosis of Hypertrophic Lichen Planus600WPMPONo ratings yet
- Send Me An AngelDocument3 pagesSend Me An AngeldeezersamNo ratings yet
- Seminars - 09-12-2022 - Vanessa AQUINO CHAVESDocument3 pagesSeminars - 09-12-2022 - Vanessa AQUINO CHAVESVanessa AquinoNo ratings yet
- Prestressed ConcreteDocument66 pagesPrestressed ConcreteTech BisuNo ratings yet
- Horgolás Minta - PulcsiDocument5 pagesHorgolás Minta - PulcsiCagey Ice-RoyNo ratings yet
- Peer-to-Peer Lending Using BlockchainDocument22 pagesPeer-to-Peer Lending Using BlockchainLuis QuevedoNo ratings yet
- Powerpoints 4 4up8Document9 pagesPowerpoints 4 4up8Ali KalyarNo ratings yet
- GMAT Sentence Correction Practice Test 03Document5 pagesGMAT Sentence Correction Practice Test 03krishnachivukulaNo ratings yet
- Tutorial Class 4: Finders As Bailee Right of A Bailee General LienDocument26 pagesTutorial Class 4: Finders As Bailee Right of A Bailee General Lienchirag jainNo ratings yet
- Answer The Statements Below by Boxing The Past Perfect TenseDocument1 pageAnswer The Statements Below by Boxing The Past Perfect TenseMa. Myla PoliquitNo ratings yet
- IN THE BEGINNING WAS AFFECT RolnikDocument22 pagesIN THE BEGINNING WAS AFFECT RolnikFabiana PaulinoNo ratings yet
- Novi Hervianti Putri - A1E015047Document2 pagesNovi Hervianti Putri - A1E015047Novi Hervianti PutriNo ratings yet
- HP MSM775 ZL Controller Installation GuideDocument21 pagesHP MSM775 ZL Controller Installation GuidezarandijaNo ratings yet
- dm00226326 Using The Hardware Realtime Clock RTC and The Tamper Management Unit Tamp With stm32 MicrocontrollersDocument82 pagesdm00226326 Using The Hardware Realtime Clock RTC and The Tamper Management Unit Tamp With stm32 MicrocontrollersMoutaz KhaterNo ratings yet
- Seven Seas of CommunicationDocument2 pagesSeven Seas of Communicationraaaj500501No ratings yet