You might also like
- Lab Report Group 6 Nerves and ReflexesDocument5 pagesLab Report Group 6 Nerves and ReflexesKristina Esta50% (2)
- Chap 7Document13 pagesChap 7VishalNo ratings yet
- Control and Coordination - Class XDocument5 pagesControl and Coordination - Class XMonika Mehan89% (19)
- Reflexes:: DefinitionDocument11 pagesReflexes:: DefinitionUMAIR JAMEELNo ratings yet
- 7.reflex Regulation of Physiological Function PDFDocument2 pages7.reflex Regulation of Physiological Function PDFNektarios TsakalosNo ratings yet
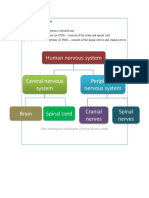
- Chart Showing The Classification of Human Nervous SystemDocument6 pagesChart Showing The Classification of Human Nervous Systemamala raviNo ratings yet
- Demonstration of The Reflex Pathway in FrogsDocument12 pagesDemonstration of The Reflex Pathway in FrogsRuwiniRajakarunaNo ratings yet
- Control and Coordination ExplainedDocument11 pagesControl and Coordination ExplainedJit AggNo ratings yet
- Biology 12 Section Assignment 4.1 Part A: Reflex Ares Laboratory Lab Report Willis Lee HypothesisDocument5 pagesBiology 12 Section Assignment 4.1 Part A: Reflex Ares Laboratory Lab Report Willis Lee HypothesisJames JungNo ratings yet
- Reflexes: Laboratory Practical General ObjectivesDocument4 pagesReflexes: Laboratory Practical General ObjectivesPhysio08No ratings yet
- Reflex MovementDocument24 pagesReflex MovementMuqeet76No ratings yet
- Reflex ArcDocument8 pagesReflex ArcHina AsifNo ratings yet
- Bio 122 FR 11 - Reflex Action in FrogsDocument8 pagesBio 122 FR 11 - Reflex Action in FrogsLance CarandangNo ratings yet
- LAPORAN ANFISMAN - Unit 4Document14 pagesLAPORAN ANFISMAN - Unit 4Rezki Asya NurNo ratings yet
- Nervous System Reflexes and Senses: Physiology Lab-4 October, 2018Document21 pagesNervous System Reflexes and Senses: Physiology Lab-4 October, 2018Madhu LodhiNo ratings yet
- Reflexes Kine Shehroz.Document12 pagesReflexes Kine Shehroz.UMAIR JAMEELNo ratings yet
- Control Coordination Nervous SystemDocument19 pagesControl Coordination Nervous SystemVanitha raoNo ratings yet
- Spinal ReflexDocument10 pagesSpinal ReflexFata HalaniNo ratings yet
- Lab 03 Reflexes Referred PainDocument14 pagesLab 03 Reflexes Referred PainViraj TatikondaNo ratings yet
- DATE: - Experiment: 01 ObjectsDocument8 pagesDATE: - Experiment: 01 ObjectsMuhammad BashirNo ratings yet
- Laprak 1 FixDocument11 pagesLaprak 1 FixTakery Chiko KeikoNo ratings yet
- Human anatomy and physiology report on reflexesDocument8 pagesHuman anatomy and physiology report on reflexesNur Fahmi UtamiNo ratings yet
- Lab - Nerve ReflexesDocument6 pagesLab - Nerve Reflexesirfan100% (1)
- Protection of the Eye: How the Eyeball, Eyelids, Tears and Other Structures Safeguard VisionDocument19 pagesProtection of the Eye: How the Eyeball, Eyelids, Tears and Other Structures Safeguard VisionReynaldo ThomasNo ratings yet
- Chap 7 Class X Cbse Science Biology.Document13 pagesChap 7 Class X Cbse Science Biology.Balqis SalamaNo ratings yet
- Human Nervous SystemDocument6 pagesHuman Nervous SystemI Kadek Adi Putra Suandana 2005No ratings yet
- Anatomical Characteristics of The Vestibular SystemDocument7 pagesAnatomical Characteristics of The Vestibular SystemRuth AlooNo ratings yet
- Refleks Spinal (Revised)Document11 pagesRefleks Spinal (Revised)Anang Yanuar RamadhanNo ratings yet
- Reflexes and Reaction Times: Student HandoutDocument7 pagesReflexes and Reaction Times: Student HandoutRema AkilNo ratings yet
- Anatomy Nervous SystemDocument13 pagesAnatomy Nervous SystemDewa NurjaNo ratings yet
- Full Body Reflexology Chart Template Free PDF Format PDFDocument37 pagesFull Body Reflexology Chart Template Free PDF Format PDFShadrina Al Shafama100% (3)
- Role of nervous system in shaping animal behaviourDocument2 pagesRole of nervous system in shaping animal behaviourMisbah NoreenNo ratings yet
- BIO 361 Neural Integration LabDocument5 pagesBIO 361 Neural Integration LabnwandspNo ratings yet
- Postural Reflex Final 2Document27 pagesPostural Reflex Final 2Shirish RaoNo ratings yet
- Exercise 13 Nervous Responses Postlab ReportDocument6 pagesExercise 13 Nervous Responses Postlab ReportAmanda SusulinNo ratings yet
- Notes On Coordination - Response - Nervous System, Reflex Arc, Type of NeuronesDocument6 pagesNotes On Coordination - Response - Nervous System, Reflex Arc, Type of NeuronesamberNo ratings yet
- The Posture & Righting Reflexes (Guyton Ed. 13: Pg. 704-705)Document21 pagesThe Posture & Righting Reflexes (Guyton Ed. 13: Pg. 704-705)Mudassar SattarNo ratings yet
- Tulip Series English GuideDocument11 pagesTulip Series English GuideUrwah hassanNo ratings yet
- ReflexDocument2 pagesReflexSarah LewisNo ratings yet
- Double-Pithed FrogDocument3 pagesDouble-Pithed FrogDeasserei TatelNo ratings yet
- Control & CoordinationDocument71 pagesControl & CoordinationPrabhnoor kaurNo ratings yet
- Spinal FrogDocument9 pagesSpinal FrogLutfia NirwanaNo ratings yet
- Neuro 1Document26 pagesNeuro 1Iris BakerNo ratings yet
- Chapter 2Document35 pagesChapter 2Amiey SehokNo ratings yet
- Reflexes Guide: 20+ Human Reflexes ExplainedDocument37 pagesReflexes Guide: 20+ Human Reflexes ExplainedSalman KhanNo ratings yet
- The Cerebelum 2Document53 pagesThe Cerebelum 2Reagan MwesigwaNo ratings yet
- Physio Psy Module 8Document4 pagesPhysio Psy Module 8Kier Elizalde VistalNo ratings yet
- Control and Coordination CL XDocument7 pagesControl and Coordination CL XMukul BanoulaNo ratings yet
- Reflexes: Yara George AbdouDocument7 pagesReflexes: Yara George AbdouVivek VijayendraNo ratings yet
- NervoussystemlabreportDocument9 pagesNervoussystemlabreportapi-296594736No ratings yet
- Jesc107 PDFDocument13 pagesJesc107 PDFpraxis TDLNo ratings yet
- Control and CordinateDocument12 pagesControl and CordinateHarsh RathoreNo ratings yet
- Von Holst 1954 Relations Between CNS and Peripheral OrgansDocument6 pagesVon Holst 1954 Relations Between CNS and Peripheral Organsrimon.redaNo ratings yet
- Vagus Nerve: A Complete Guide to Activate the Healing power of Your Vagus Nerve – Reduce with Self-Help Exercises Anxiety, PTSD, Chronic Illness, Depression, Inflammation, Anger and TraumaFrom EverandVagus Nerve: A Complete Guide to Activate the Healing power of Your Vagus Nerve – Reduce with Self-Help Exercises Anxiety, PTSD, Chronic Illness, Depression, Inflammation, Anger and TraumaNo ratings yet
- Exercise 7.2 - ElectrocardiogramDocument2 pagesExercise 7.2 - ElectrocardiogramKevin F. CortesNo ratings yet
- Rabbit Breeds Management and ClassificationDocument37 pagesRabbit Breeds Management and ClassificationAÿâñ Jitéñdrä Yãdáv100% (1)
- Finding An Apartment Project: Background - Answer These Questions in Complete SentencesDocument4 pagesFinding An Apartment Project: Background - Answer These Questions in Complete Sentencesapi-288310744No ratings yet
- Histology of Small Intestine 3Document17 pagesHistology of Small Intestine 3ShizraNo ratings yet
- n6 Andrzej Sapkowski Season - of - StormsDocument9 pagesn6 Andrzej Sapkowski Season - of - StormsRazvan Visoiu0% (2)
- Vertigo and ImbalanceDocument19 pagesVertigo and ImbalanceDiena HarisahNo ratings yet
- Grade 9 Sex Ed - Youtube LinksDocument16 pagesGrade 9 Sex Ed - Youtube Linksapi-204719782No ratings yet
- OSCE Mock ExamDocument52 pagesOSCE Mock Examanas100% (2)
- Motor Neurone Disease and HomoeopathyDocument8 pagesMotor Neurone Disease and HomoeopathyDr. Rajneesh Kumar Sharma MD HomNo ratings yet
- Reproductive SystemDocument7 pagesReproductive SystemRiaz siddiquiNo ratings yet
- Normal Puerperium PDFDocument22 pagesNormal Puerperium PDFwawa3385100% (1)
- The Hall of Fire 05Document29 pagesThe Hall of Fire 05maldreidor100% (1)
- Bone Marrow Morphology: Normal and Abnormal FindingsDocument51 pagesBone Marrow Morphology: Normal and Abnormal FindingsMohammed FareedNo ratings yet
- Diverticular Disease 2Document5 pagesDiverticular Disease 2karl abiaadNo ratings yet
- Erythropoiesis: Presented by DR Etu-Efeotor T. PDocument88 pagesErythropoiesis: Presented by DR Etu-Efeotor T. PPrincewill SeiyefaNo ratings yet
- Grabovoi NumbersDocument29 pagesGrabovoi Numberssillyminds100% (3)
- Anatomy and Physiology of The Thyroid and Parathyroid GlandsDocument14 pagesAnatomy and Physiology of The Thyroid and Parathyroid GlandsSidiq AboobakerNo ratings yet
- Chapter 8 MariebDocument5 pagesChapter 8 MariebmjhenkeNo ratings yet
- Routine Immunization - Details - For 4th August MetingDocument57 pagesRoutine Immunization - Details - For 4th August Metingjalanayush777No ratings yet
- Statement From Maine Veterinary Medical CenterDocument3 pagesStatement From Maine Veterinary Medical CenterWGMENo ratings yet
- Chiari CaseDocument27 pagesChiari CaseShaurya AgarwalNo ratings yet
- BIOLOGY RevisionNotes SaveMyExamsDocument7 pagesBIOLOGY RevisionNotes SaveMyExamsAli AshrafNo ratings yet
- Upper Midline CorrectionDocument5 pagesUpper Midline CorrectionmutansNo ratings yet
- Toshihiko Yayama - Qi Healing - The Way To A New Mind and BodyDocument159 pagesToshihiko Yayama - Qi Healing - The Way To A New Mind and BodyFernando Iglesias100% (2)
- Vital Spots 10 - Taranaki ITF Taekwondo PDFDocument54 pagesVital Spots 10 - Taranaki ITF Taekwondo PDFMaria Elisa MonclusNo ratings yet
- CannulaDocument4 pagesCannulaAnonymous m0vrFqAsNo ratings yet
- Body Fluid Electrolytes Acid Base BalanceDocument2 pagesBody Fluid Electrolytes Acid Base BalanceMarzanul Islam AnikNo ratings yet
- A Case PresentationDocument68 pagesA Case PresentationMaxGabilanNo ratings yet
- Polio OutbreakDocument3 pagesPolio OutbreakAyn Jel MayNo ratings yet
- Conduct of Normal Labor and DeliveryDocument29 pagesConduct of Normal Labor and DeliveryEzekiel ArtetaNo ratings yet