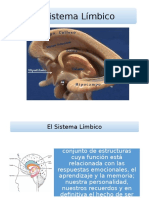
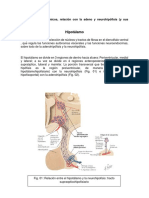
You might also like
- Sistema Nervioso SimpáticoDocument27 pagesSistema Nervioso SimpáticoGiovaniAvendañoGarciaNo ratings yet
- Sensibilidades Somáticas I. Cap. 47 GuytonDocument24 pagesSensibilidades Somáticas I. Cap. 47 GuytonVictor Eloy Gonzalez50% (6)
- Los Órganos de Los SentidosDocument13 pagesLos Órganos de Los Sentidosdolores ecijaNo ratings yet
- Funciones Reproductoras y Hormonales Masculinas (Y FunciónDocument20 pagesFunciones Reproductoras y Hormonales Masculinas (Y FunciónJoão Victor OliveiraNo ratings yet
- Ensayo OlfatoDocument4 pagesEnsayo OlfatoJulio Cesar MartimezNo ratings yet
- Pares Craneales y Su FuncionDocument25 pagesPares Craneales y Su FuncionRobinsonGiraldoLópez100% (1)
- Sistema Limbico PresentaciónDocument33 pagesSistema Limbico PresentaciónSaraDiazRueda100% (2)
- Los Sentidos EspecialesDocument25 pagesLos Sentidos Especialessulin daniela puerto ovando100% (1)
- Sistema Nervioso Simpático y ParasimpaticoDocument5 pagesSistema Nervioso Simpático y ParasimpaticoMarcelo Soledispa100% (1)
- Funcion Del Sentido Del GustoDocument3 pagesFuncion Del Sentido Del GustofrancysNo ratings yet
- El Sistema Nervioso Autónomo y La Médula SuprarrenalDocument3 pagesEl Sistema Nervioso Autónomo y La Médula SuprarrenalPablo Soto OsunaNo ratings yet
- Fisiología Sensorial 6Document54 pagesFisiología Sensorial 6Geraldine MsvNo ratings yet
- Clase de Encefalo III Sistema Limbico (p2)Document47 pagesClase de Encefalo III Sistema Limbico (p2)diliana sotoNo ratings yet
- Nociones Básicas Del Sistema NerviosoDocument16 pagesNociones Básicas Del Sistema NerviosoAndres FernandoNo ratings yet
- MeningesDocument24 pagesMeningesRijardNo ratings yet
- Principios Generales y Fisiologia SensorialDocument24 pagesPrincipios Generales y Fisiologia SensorialCarlos Javier MeroNo ratings yet
- Función de Las Glándulas EndocrinasDocument5 pagesFunción de Las Glándulas EndocrinasAlvaro CastilloNo ratings yet
- Pares Craneales o Nervios CranealesDocument10 pagesPares Craneales o Nervios CranealesAntonio Arroyo CornejoNo ratings yet
- Trabajo Pares CranealesDocument54 pagesTrabajo Pares CranealesLuulaa Minguin GarciaNo ratings yet
- Sentidos EspecialesDocument8 pagesSentidos EspecialesIon Frias0% (1)
- Exploración Por RegionesDocument33 pagesExploración Por RegionesfannyfanssNo ratings yet
- Cuestionario RespiraciónDocument5 pagesCuestionario RespiraciónGonzalo Molina GonzalezNo ratings yet
- EstertoresDocument7 pagesEstertoresFernando Sánchez DíezNo ratings yet
- Clase 1 Sensación y PercepciónDocument13 pagesClase 1 Sensación y PercepciónkryssemmaNo ratings yet
- Sistema Nervioso Sensorial.: Aprendizaje Basado en ProblemasDocument38 pagesSistema Nervioso Sensorial.: Aprendizaje Basado en ProblemasAna Cristina AlvarezNo ratings yet
- Corteza Cerebral, Funciones Intelectuales Del Cerebro, Sensaciones Táctil y PosicionalDocument103 pagesCorteza Cerebral, Funciones Intelectuales Del Cerebro, Sensaciones Táctil y PosicionaldiowhgblNo ratings yet
- HDocument10 pagesHGeorge BetancourthNo ratings yet
- Control de La Función Motora Por La Corteza y El Tronco EncefálicoDocument7 pagesControl de La Función Motora Por La Corteza y El Tronco EncefálicoCésar Malvaez100% (1)
- Psicopatologia ForenseDocument30 pagesPsicopatologia ForenseSara Ortega PhotographyNo ratings yet
- Nervio Vestibulococlear, AnatomíaDocument3 pagesNervio Vestibulococlear, AnatomíaMartín MeléndezNo ratings yet
- Tronco EncefálicoDocument21 pagesTronco EncefálicoJavier VelasquezNo ratings yet
- Sistema Sensorial VisualDocument3 pagesSistema Sensorial VisualYoseth HernandezNo ratings yet
- Formaciones Interhemisféricas 2Document30 pagesFormaciones Interhemisféricas 2mirka67% (3)
- Potencial de MembranaDocument15 pagesPotencial de MembranaScarlethMuñoz100% (1)
- 3 y 4 HipofisisDocument6 pages3 y 4 HipofisisAníbal CINo ratings yet
- Escuelas PsicológicasDocument2 pagesEscuelas PsicológicasLoty Vergara VasquezNo ratings yet
- (9E) Tegumento - Miología - HombroDocument35 pages(9E) Tegumento - Miología - HombroNayely GVNo ratings yet
- 3.ppse y PpsiDocument63 pages3.ppse y PpsiJOhn JacoboNo ratings yet
- Sistema Cardiovascular Practica - HechaDocument20 pagesSistema Cardiovascular Practica - HechaJean Carlos Rivera AlmonteNo ratings yet
- Trastornos Relacionados Con Sedantes HipnóticosDocument31 pagesTrastornos Relacionados Con Sedantes HipnóticosBeatriz MarquezNo ratings yet
- HORMONADocument3 pagesHORMONANelida Gonzalez BaezNo ratings yet
- Ventilacion-Pulmonar MonografiaDocument7 pagesVentilacion-Pulmonar MonografiaHellen Mallaupoma ObregonNo ratings yet
- La MelatoninaDocument2 pagesLa MelatoninaPaty Mend100% (1)
- Sentidos EspecialesDocument22 pagesSentidos Especialesgenesis100% (1)
- Sueño y VigiliaDocument34 pagesSueño y VigiliaLuis Aquino GagoNo ratings yet
- Las Pseudoalucinaciones SEMAS 6Document3 pagesLas Pseudoalucinaciones SEMAS 6Ivettebrenda HualpaNo ratings yet
- Fisiología Del Sentido Del Tacto y Audición IIDocument11 pagesFisiología Del Sentido Del Tacto y Audición IILety CatariNo ratings yet
- Pares Craneales FlashcardsDocument6 pagesPares Craneales FlashcardsMaría José Hernández LunaNo ratings yet
- Patologia Del HipotalamoDocument10 pagesPatologia Del HipotalamoPaola Del BarrioNo ratings yet
- 11.sistema EndocrinoDocument47 pages11.sistema Endocrinojeniffer YmNo ratings yet
- Resumen 1er Capítulo de SnellDocument4 pagesResumen 1er Capítulo de SnellSalma MendozaNo ratings yet
- 03 - Alteraciones de La ConcienciaDocument34 pages03 - Alteraciones de La ConcienciaAle ZambranoNo ratings yet
- Los Sentidos QuímicosDocument35 pagesLos Sentidos QuímicosSandy MendNo ratings yet
- Cuestionario de SBV (2 Parcial)Document5 pagesCuestionario de SBV (2 Parcial)Roy Fernando Molina PaccoNo ratings yet
- Codigo de NurembergDocument2 pagesCodigo de NurembergJhon Robert Ramirez100% (2)
- Nacimiento y Primeros Desarrollos de La Psicología Científica en Las Islas BritánicasDocument2 pagesNacimiento y Primeros Desarrollos de La Psicología Científica en Las Islas BritánicasThomas GonzalezNo ratings yet
- Masoterapia en Las Alteraciones Conductuales de Ancianos Con DemenciaDocument2 pagesMasoterapia en Las Alteraciones Conductuales de Ancianos Con DemenciaFrancisco AriasNo ratings yet
- Ensayo 2Document13 pagesEnsayo 2Guillermo PreciadoNo ratings yet
- 48Document5 pages48Jhony PradelaNo ratings yet
- Sensibilidades Somaticas 1Document9 pagesSensibilidades Somaticas 1MARIANA AILEN FLORES FLORESNo ratings yet
- Hoja Recomendaciones EDADocument2 pagesHoja Recomendaciones EDArodrigo4590No ratings yet
- LeucemiasDocument4 pagesLeucemiasrodrigo4590No ratings yet
- Solicitud de Exámenes de LaboratorioDocument2 pagesSolicitud de Exámenes de Laboratoriorodrigo4590No ratings yet
- Análisis de Artículo PRISMADocument3 pagesAnálisis de Artículo PRISMArodrigo4590No ratings yet
- Trastornos Hematológicos Del Recién NacidoDocument30 pagesTrastornos Hematológicos Del Recién Nacidorodrigo4590100% (1)
- Cap 7 Cartílago y HuesoDocument18 pagesCap 7 Cartílago y Huesorodrigo4590No ratings yet
- Declaración CONSORTDocument2 pagesDeclaración CONSORTrodrigo4590No ratings yet
- Cap 10 Sangre y HematopoyesisDocument12 pagesCap 10 Sangre y Hematopoyesisrodrigo4590No ratings yet
- Cap 13 Sistema EndócrinoDocument10 pagesCap 13 Sistema Endócrinorodrigo4590No ratings yet
- Cap 1 Introducción A La Histología y Técnicas Histológicas BásicasDocument8 pagesCap 1 Introducción A La Histología y Técnicas Histológicas Básicasrodrigo4590100% (2)
- Cap 6 Contracción Del Músculo EsqueléticoDocument9 pagesCap 6 Contracción Del Músculo Esqueléticorodrigo4590No ratings yet
- Cap 7 Excitación Del Músculo Esquelético - Transmisión Neuromuscular y Acoplamiento Excitación-ContracciónDocument4 pagesCap 7 Excitación Del Músculo Esquelético - Transmisión Neuromuscular y Acoplamiento Excitación-Contracciónrodrigo4590100% (3)
- Cap 6 Fármacos Con Acción en El Sistema Nervioso Autónomo - IntroducciónDocument4 pagesCap 6 Fármacos Con Acción en El Sistema Nervioso Autónomo - Introducciónrodrigo4590No ratings yet
- Cap 4 Biotransformación FarmacológicaDocument5 pagesCap 4 Biotransformación Farmacológicarodrigo4590No ratings yet
- Cap 8 Fármacos Antagonistas de Los ColinorreceptoresDocument9 pagesCap 8 Fármacos Antagonistas de Los Colinorreceptoresrodrigo45900% (1)
- Simulacro Residentado Medico Parte B Con ClavesDocument8 pagesSimulacro Residentado Medico Parte B Con ClavesKellyMarTegraNo ratings yet
- Cartilla27 2016Document32 pagesCartilla27 2016Antonio PazNo ratings yet
- Ficha TRACTOR M9540-CON-CABINADocument2 pagesFicha TRACTOR M9540-CON-CABINAjeissonNo ratings yet
- ABEJASDocument52 pagesABEJASHernán Castro SanabriaNo ratings yet
- La Historia de Una GaviotaDocument14 pagesLa Historia de Una GaviotaLilianaCvitanicNo ratings yet
- El HerraderoDocument8 pagesEl HerraderoMichelle Pérez GarciaNo ratings yet
- TemDocument17 pagesTemJuan TamaniNo ratings yet
- San Pedro Mártir de VeronaDocument18 pagesSan Pedro Mártir de VeronaHugoRodriguezSanMiguelNo ratings yet
- 01 Problemas de Genética I. Las Leyes de MendelDocument2 pages01 Problemas de Genética I. Las Leyes de MendelalmudenaNo ratings yet
- Cuáles Son Los Cinco Grandes Rasgos de PersonalidadDocument5 pagesCuáles Son Los Cinco Grandes Rasgos de PersonalidadtaniaNo ratings yet
- Significado de A1, ..... AaashtoDocument4 pagesSignificado de A1, ..... AaashtoFernandezMendozaBrayanValentin 5to ANo ratings yet
- 08 PPI Tabiqueria InteriorDocument1 page08 PPI Tabiqueria Interioredson eduardoNo ratings yet
- Finesse WG MSDS FMC PDFDocument7 pagesFinesse WG MSDS FMC PDFMarianoNo ratings yet
- PresbifoniaDocument25 pagesPresbifoniaCamila Paz100% (2)
- Tipologia Vehicular NTC 3729Document13 pagesTipologia Vehicular NTC 3729Carlos Barrera ReyesNo ratings yet
- Comprension de Textos TerminadoDocument16 pagesComprension de Textos Terminadoanon_8411093690% (1)
- Resumen Unidad III2Document23 pagesResumen Unidad III2Italo AdottiNo ratings yet
- Gestalt para PrincipiantesDocument4 pagesGestalt para Principiantesluisa diazarNo ratings yet
- Regresion Logic en SPSSDocument16 pagesRegresion Logic en SPSSeconomistasdelawebNo ratings yet
- FARMACODINAMIADocument34 pagesFARMACODINAMIAArturo Esquivel100% (2)
- Procesos AuxiliaresDocument30 pagesProcesos AuxiliaresJosé Manuel M.L.No ratings yet
- Examen PuentesDocument15 pagesExamen PuentesCarlos Daniel Sanchez MejiaNo ratings yet
- PioglitazonaDocument2 pagesPioglitazonaCarlosMarquezPNo ratings yet
- TRABAJO PRCTICO DE PROCEDIMIENTOS BASICOS DE SALUD Semana 7Document19 pagesTRABAJO PRCTICO DE PROCEDIMIENTOS BASICOS DE SALUD Semana 7rafael moreta floresNo ratings yet
- Revista Electrónica de AnestesiarDocument6 pagesRevista Electrónica de AnestesiarVIVIAN DANIEL SUPELANO MILLANNo ratings yet
- Vocabulario Sobre MaquinasDocument4 pagesVocabulario Sobre MaquinasSandy Yoselinne Samudio GaitanNo ratings yet
- Remodelacion e Implementacion de LabortorioDocument6 pagesRemodelacion e Implementacion de LabortorioGuillermo Gutierrez50% (2)
- Unidad V Dibujos de DefiniciónDocument20 pagesUnidad V Dibujos de DefiniciónKARLA ALARCONNo ratings yet
- Astronomia - Practicas de Geologia PlanetariaDocument8 pagesAstronomia - Practicas de Geologia PlanetariaFrancisco Javier Sánchez CruzNo ratings yet