You might also like
- Principios de Fisiología SensorialDocument21 pagesPrincipios de Fisiología SensorialVictor Choque FloresNo ratings yet
- Caso clínico de ansiedad socialDocument6 pagesCaso clínico de ansiedad socialsilviaNo ratings yet
- Adulto JovenDocument19 pagesAdulto JovenzulemaNo ratings yet
- TermorregulaciónDocument9 pagesTermorregulaciónEmmy OlsNo ratings yet
- Principios Generales y Fisiologia SensorialDocument24 pagesPrincipios Generales y Fisiologia SensorialCarlos Javier MeroNo ratings yet
- Regulacion de La Temperatura CorporalDocument25 pagesRegulacion de La Temperatura CorporalDouglas Menjivar100% (1)
- Control Del Flujo SanguíneoDocument16 pagesControl Del Flujo SanguíneoJavier Manuel Escobedo CalderónNo ratings yet
- Monografia de Etica ClínicaDocument19 pagesMonografia de Etica ClínicaSandra Liz Choqueneira FloresNo ratings yet
- Modelos salud: biomédico vs biopsicosocialDocument1 pageModelos salud: biomédico vs biopsicosocialShalom AlejemNo ratings yet
- BulimiaDocument22 pagesBulimiaAngie Velez SancanNo ratings yet
- Histologia - Sistema Endocrino AcbDocument78 pagesHistologia - Sistema Endocrino AcbMiguel Angel Mendoza100% (1)
- Exploración de Cuello y TiroidesDocument12 pagesExploración de Cuello y TiroidesJhony ArzacNo ratings yet
- Historia de La Enfermedad de WolmanDocument2 pagesHistoria de La Enfermedad de Wolmancepj1908No ratings yet
- ENDOCRINOLOGADocument36 pagesENDOCRINOLOGAanalizitap100% (1)
- Psicopatologia Inteligencia y MemoriaDocument31 pagesPsicopatologia Inteligencia y MemoriaJosueAvalosChipanaNo ratings yet
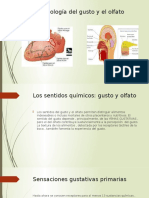
- Fisiología del gusto y olfatoDocument53 pagesFisiología del gusto y olfatoambar100% (2)
- Motilidad GastrointestinalDocument1 pageMotilidad GastrointestinalAri Santiago0% (1)
- Psicologia Medica CLASE 1Document33 pagesPsicologia Medica CLASE 1Roberto Castillo SuarezNo ratings yet
- Sistema Neuroendocrino-Diapositivas (Modo de Compatibilidad)Document3 pagesSistema Neuroendocrino-Diapositivas (Modo de Compatibilidad)Siegfried WolfteamNo ratings yet
- Farmacocinetica y FarmacodinamiaDocument21 pagesFarmacocinetica y FarmacodinamiaShaun Smith100% (1)
- (A) La Etica Filosofica y La BioeticaDocument10 pages(A) La Etica Filosofica y La BioeticaRobertoNo ratings yet
- Sospecha de Pubertad PrecozDocument20 pagesSospecha de Pubertad PrecozInmaculada Concepcion Vazquez CanoNo ratings yet
- Corteza CerebralDocument51 pagesCorteza CerebralJaime SolarteNo ratings yet
- Lectura 1 Conceptos e IndicadoresDocument15 pagesLectura 1 Conceptos e IndicadoresasesprodusoftNo ratings yet
- PP Hipotalamo Hipofisis HormonasDocument54 pagesPP Hipotalamo Hipofisis HormonasHernán FernándezNo ratings yet
- Neoplasias OvaricasDocument47 pagesNeoplasias OvaricasFrancisco JuárezNo ratings yet
- Farmacología Sistema NerviosoDocument45 pagesFarmacología Sistema NerviosocatalinaNo ratings yet
- Hormonas y conducta sexualDocument19 pagesHormonas y conducta sexualHugoNo ratings yet
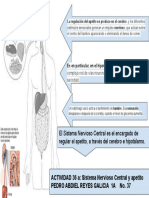
- Esquema SNC y Apetito PDFDocument1 pageEsquema SNC y Apetito PDFPedroReyesLopezNo ratings yet
- Pares Craneales-1Document24 pagesPares Craneales-1Jean BrenesNo ratings yet
- CITOCINASDocument11 pagesCITOCINASMaría Camila SánchezNo ratings yet
- Signos y Síntomas UrinariosDocument13 pagesSignos y Síntomas UrinariosL Fernando TordoyaNo ratings yet
- Dss Sostenible - OdsDocument70 pagesDss Sostenible - OdsFrancisco A Risso ANo ratings yet
- Hepáticas, Vesícula, Bazo y PáncreasDocument30 pagesHepáticas, Vesícula, Bazo y PáncreasEsteban MorochoNo ratings yet
- Capítulo 1. Introducción A La FarmacologíaDocument9 pagesCapítulo 1. Introducción A La FarmacologíaAlexandra ZarateNo ratings yet
- Periodo FetalDocument9 pagesPeriodo FetalFlor Chumacero ZegarraNo ratings yet
- Sistema Nervioso Periferico - FisiologíaDocument6 pagesSistema Nervioso Periferico - FisiologíaPadron JoseNo ratings yet
- Sentidos EspecialesDocument22 pagesSentidos Especialesgenesis100% (1)
- Cuadro Comparativo Neuronas (Autoguardado)Document2 pagesCuadro Comparativo Neuronas (Autoguardado)dianavictoria lozada100% (1)
- Sistema Endocrino Universidad Mariano Gálvez GuatemalaDocument12 pagesSistema Endocrino Universidad Mariano Gálvez GuatemalaLuis Felipe MazariegosNo ratings yet
- CalcitoninaDocument2 pagesCalcitoninaRodríguez Lovera Juan CarlosNo ratings yet
- Valoracion de Mamas y AxilasDocument30 pagesValoracion de Mamas y AxilasDaniela ContrerasNo ratings yet
- Introduccion V JSTDocument62 pagesIntroduccion V JSTMEDICINA UAEMNo ratings yet
- Informe de Laboratorio # 3 Sensibiliad - Gusto y OlfatoDocument10 pagesInforme de Laboratorio # 3 Sensibiliad - Gusto y OlfatoCamilo Salinas100% (1)
- La Bioetica y Las AdiccionesDocument3 pagesLa Bioetica y Las AdiccionesTonanzin Guadalupe Olivera Molina100% (1)
- Mapa Conceptual - Psicologia HumanistaDocument1 pageMapa Conceptual - Psicologia HumanistaBP OyukiNo ratings yet
- Vía Piramidal - Perez Elena, Perez Deyni, Perez Anavel, Portilla EstefhaniDocument13 pagesVía Piramidal - Perez Elena, Perez Deyni, Perez Anavel, Portilla EstefhaniJHON JHERICO DOMINGUEZ CORDOVA100% (1)
- Ex Posicion EsDocument186 pagesEx Posicion EsJonathan DominguezNo ratings yet
- Tecnicas de Exploracion Fisica - MVG - Julio.2018Document46 pagesTecnicas de Exploracion Fisica - MVG - Julio.2018Cesar Del Aguila Matta100% (2)
- Metodología PostestructuralismoDocument46 pagesMetodología PostestructuralismoCarla Almeida50% (2)
- Coagulopatias AdquiridasDocument9 pagesCoagulopatias AdquiridasEdwar Guido Briceño EsquivelNo ratings yet
- HongosDocument58 pagesHongosDaniel perez perez100% (1)
- Funciones Secretoras Del Tubo Digestivo-Grupo 3Document33 pagesFunciones Secretoras Del Tubo Digestivo-Grupo 3Emily ZambranoNo ratings yet
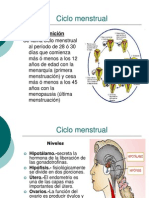
- Ciclo menstrual: fases, períodos y vías de administración de medicamentosDocument18 pagesCiclo menstrual: fases, períodos y vías de administración de medicamentosDelia Eliana Chura Totora0% (1)
- Haemophilus ducreyi: agente causal del chancro blandoDocument1 pageHaemophilus ducreyi: agente causal del chancro blandoingrit vazquezNo ratings yet
- Evaluacion PsicologicaDocument32 pagesEvaluacion Psicologica05-ES-HU-RAY CHARLIE CARHUAMACA ORTIZNo ratings yet
- LaSaludPublicaYElTrabajoEnComunidad GonzalezDocument188 pagesLaSaludPublicaYElTrabajoEnComunidad GonzalezGisel CasanovaNo ratings yet
- Resumen Tercera Semana de DesarrolloDocument3 pagesResumen Tercera Semana de DesarrolloAugusto Crocker0% (1)
- Receptores sensoriales y procesamiento neuronalDocument8 pagesReceptores sensoriales y procesamiento neuronalKony Ramos RojasNo ratings yet
- Guía Expocision FisioDocument8 pagesGuía Expocision FisioLuz Dary Orbes ConstainNo ratings yet
- Hoja Recomendaciones EDADocument2 pagesHoja Recomendaciones EDArodrigo4590No ratings yet
- Solicitud de Exámenes de LaboratorioDocument2 pagesSolicitud de Exámenes de Laboratoriorodrigo4590No ratings yet
- LeucemiasDocument4 pagesLeucemiasrodrigo4590No ratings yet
- Análisis de Artículo PRISMADocument3 pagesAnálisis de Artículo PRISMArodrigo4590No ratings yet
- GlosarioDocument1 pageGlosariorodrigo4590No ratings yet
- Trastornos Hematológicos Del Recién NacidoDocument30 pagesTrastornos Hematológicos Del Recién Nacidorodrigo4590100% (1)
- ChecoDocument18 pagesChecorodrigo4590No ratings yet
- Análisis de ArtículoDocument8 pagesAnálisis de Artículorodrigo4590No ratings yet
- Atención Primaria de La SaludDocument2 pagesAtención Primaria de La Saludrodrigo4590No ratings yet
- Análisis de DecisionesDocument16 pagesAnálisis de Decisionesrodrigo4590No ratings yet
- Ascitis y EsputoDocument17 pagesAscitis y Esputorodrigo4590No ratings yet
- DefinicionesDocument4 pagesDefinicionesrodrigo4590No ratings yet
- Cap 7 Cartílago y HuesoDocument18 pagesCap 7 Cartílago y Huesorodrigo4590No ratings yet
- Casos ClínicosDocument7 pagesCasos Clínicosrodrigo4590No ratings yet
- CefaleasDocument6 pagesCefaleasrodrigo4590No ratings yet
- Cap 3 NúcleoDocument9 pagesCap 3 Núcleorodrigo4590No ratings yet
- Estadística InferencialDocument16 pagesEstadística Inferencialrodrigo4590No ratings yet
- Declaración CONSORTDocument2 pagesDeclaración CONSORTrodrigo4590No ratings yet
- Cap 11 Sistema CirculatorioDocument7 pagesCap 11 Sistema Circulatoriorodrigo4590No ratings yet
- Cap 5 Epitelio y GlándulasDocument14 pagesCap 5 Epitelio y Glándulasrodrigo4590No ratings yet
- Cap 1 Introducción A La Histología y Técnicas Histológicas BásicasDocument8 pagesCap 1 Introducción A La Histología y Técnicas Histológicas Básicasrodrigo4590100% (2)
- Análisis de SobrevidaDocument1 pageAnálisis de Sobrevidarodrigo4590No ratings yet
- Cap 6 Tejido ConectivoDocument10 pagesCap 6 Tejido Conectivorodrigo4590100% (1)
- Cap 4 Matriz ExtracelularDocument5 pagesCap 4 Matriz Extracelularrodrigo4590No ratings yet
- Cap 13 Sistema EndócrinoDocument10 pagesCap 13 Sistema Endócrinorodrigo4590No ratings yet
- Sistema nervioso, sinapsis y neurotransmisoresDocument11 pagesSistema nervioso, sinapsis y neurotransmisoresrodrigo4590100% (2)
- Cap 10 Sangre y HematopoyesisDocument12 pagesCap 10 Sangre y Hematopoyesisrodrigo4590No ratings yet
- Cap 6 Contracción Del Músculo EsqueléticoDocument9 pagesCap 6 Contracción Del Músculo Esqueléticorodrigo4590No ratings yet
- Cap 46 Receptores Sensitivos, Circuitos Neuronales para El Procesamiento de La InformaciónDocument8 pagesCap 46 Receptores Sensitivos, Circuitos Neuronales para El Procesamiento de La Informaciónrodrigo4590No ratings yet
- Sensibilidad somática: receptores táctiles y vías de transmisiónDocument8 pagesSensibilidad somática: receptores táctiles y vías de transmisiónrodrigo4590100% (1)
- Formato Guia de Autoaprendizaje Guía 9-IDocument14 pagesFormato Guia de Autoaprendizaje Guía 9-IAmatapi ApellidoNo ratings yet
- Cortico Ponto CerebelosaDocument4 pagesCortico Ponto CerebelosaAngélica RomanNo ratings yet
- NeuroanatomíaV PDFDocument431 pagesNeuroanatomíaV PDFRonald Jarl Bandam100% (3)
- TeaDocument6 pagesTeaCarolina Saez PachecoNo ratings yet
- Programa Nivel 1 en PFD NNASCDocument67 pagesPrograma Nivel 1 en PFD NNASCfabricioio gumiel forenzaNo ratings yet
- DISECCION DEL ENCEFALO Lab N°3Document9 pagesDISECCION DEL ENCEFALO Lab N°3yeissy perezsNo ratings yet
- Trabajo Final Neuropsicologia El CerebeloDocument14 pagesTrabajo Final Neuropsicologia El CerebeloOsvaldo vasquezNo ratings yet
- Eco Cerebral: Indicaciones, Características y CortesDocument15 pagesEco Cerebral: Indicaciones, Características y CortesperuxaNo ratings yet
- TFG CD14 34356 Tornal Coll Andrea MJ35364 07062019Document51 pagesTFG CD14 34356 Tornal Coll Andrea MJ35364 07062019Andrea Tornal CollNo ratings yet
- Cerebro y CerebeloDocument2 pagesCerebro y CerebeloMartin Gonzalez HernandezNo ratings yet
- Cerebro Histología Estructura 40Document4 pagesCerebro Histología Estructura 40Claudio Caballero LópezNo ratings yet
- Via EspinocerebeloDocument4 pagesVia EspinocerebeloElisa CamposNo ratings yet
- 07 GAP Sistematica Neurosonografia FetalDocument22 pages07 GAP Sistematica Neurosonografia FetalAnulfo ParedesNo ratings yet
- Clase 10 TC Anatomia Normal, Protocolos de SNCDocument27 pagesClase 10 TC Anatomia Normal, Protocolos de SNCJorge Fabián Neira GamboaNo ratings yet
- CerebeloDocument4 pagesCerebelodafne de la barraNo ratings yet
- Neurociencia. Telencefalo PDFDocument388 pagesNeurociencia. Telencefalo PDFWilly Fernandez100% (8)
- Resumen - SNC IDocument29 pagesResumen - SNC IricardoNo ratings yet
- Ce RebeloDocument9 pagesCe RebeloBrems MarNo ratings yet
- Informe Medula y Tronco Encefalico-AnatomiaDocument9 pagesInforme Medula y Tronco Encefalico-Anatomiafrancis_bscNo ratings yet
- MORFOLOGÍA SISTEMA NERVIOSO - 01 Uac ValdiviaDocument30 pagesMORFOLOGÍA SISTEMA NERVIOSO - 01 Uac Valdiviavitoxis100% (10)
- Sistemas descendentes y ascendentes de la médula espinalDocument42 pagesSistemas descendentes y ascendentes de la médula espinalYuri Medrano PlanaNo ratings yet
- 2017-Cerebro, Psiquismo y SociedadDocument64 pages2017-Cerebro, Psiquismo y SociedadtipeossNo ratings yet
- 5 CerebeloDocument45 pages5 CerebeloNataNo ratings yet
- Fucs PDFDocument54 pagesFucs PDFAngélica Peña OrtizNo ratings yet
- Venas de La Cabeza y Del Cuello ArregladDocument43 pagesVenas de La Cabeza y Del Cuello ArregladYeny Apaza Calizaya0% (1)
- Bases Neurobiológicas Del Lenguaje, La Deglución y El HablaDocument25 pagesBases Neurobiológicas Del Lenguaje, La Deglución y El HablaFreddy Adam Mendez MoncadaNo ratings yet
- Cerebro Exposicion - 1Document37 pagesCerebro Exposicion - 1Silvana YogaNo ratings yet
- Neuroanatomia 182343 Downloadable 3531155Document34 pagesNeuroanatomia 182343 Downloadable 3531155hitomik00No ratings yet
- Repaso Examen Final de HistologiaDocument2 pagesRepaso Examen Final de HistologiaJennifer BrunoNo ratings yet
- Mapa Conceptual Sistema Nervioso Central y AutonomoDocument3 pagesMapa Conceptual Sistema Nervioso Central y AutonomoDianita Curimilma90% (10)