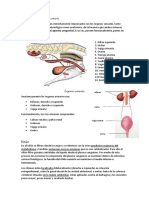
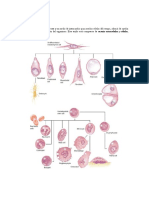
You might also like
- Circuito de PapezDocument4 pagesCircuito de PapezHans Solorzano VilchezNo ratings yet
- Corteza CerebralDocument25 pagesCorteza CerebralAlejandra Garcia ObregonNo ratings yet
- Capitulo 55 FisioDocument2 pagesCapitulo 55 FisioVickyFingadoNo ratings yet
- Nervio AisladoDocument33 pagesNervio AisladoMartha HerreraNo ratings yet
- Control de Temperatura y Nivel Con PLCDocument21 pagesControl de Temperatura y Nivel Con PLCdiegofernandodiego90% (10)
- Formación Reticular, Vigilia y SueñoDocument43 pagesFormación Reticular, Vigilia y Sueñoelinchapelotas100% (1)
- Corteza cerebral funciones intelectuales 40Document4 pagesCorteza cerebral funciones intelectuales 40Mirian GutierrezNo ratings yet
- Lo que es el subtálamoDocument4 pagesLo que es el subtálamoYuli AndradeNo ratings yet
- Sistema Nervioso SimpáticoDocument27 pagesSistema Nervioso SimpáticoGiovaniAvendañoGarciaNo ratings yet
- Capítulo 45 GuytonDocument51 pagesCapítulo 45 GuytonBillcar Calderon88% (8)
- Anatomía y Fisiología de La Glándula HipófisisDocument3 pagesAnatomía y Fisiología de La Glándula HipófisisdrawdedreamsNo ratings yet
- NeuroanatomiaDocument23 pagesNeuroanatomiaamshoteluis90% (10)
- Desarrollo Embrionario SISTEMA NERVIOSO-1Document70 pagesDesarrollo Embrionario SISTEMA NERVIOSO-1Brenda Gress ViverosNo ratings yet
- Resumen Del Capitulo 45 de Guyton (Sistema Nervioso)Document12 pagesResumen Del Capitulo 45 de Guyton (Sistema Nervioso)jorge amador100% (1)
- Embriologia Del Tronco EncefálicoDocument5 pagesEmbriologia Del Tronco EncefálicoMARIA AZUCENA CRUZADO TOLEDONo ratings yet
- Anatomia CuadroDocument4 pagesAnatomia CuadroGénesis LeónNo ratings yet
- Glosario NeurocieciasDocument23 pagesGlosario Neurocieciasslimj502No ratings yet
- L.E. Natanael Librado GonzálezDocument54 pagesL.E. Natanael Librado GonzálezNatanael LibradoNo ratings yet
- Vias OlfatoriasDocument6 pagesVias OlfatoriasSherly Angelica Vidarte Cossio100% (2)
- Anatomía Del Gusto y El OlfatoDocument16 pagesAnatomía Del Gusto y El OlfatoGeraldine Serna VasquezNo ratings yet
- Informe Tejido EpitelialDocument20 pagesInforme Tejido EpitelialAnyelo Amaya CastroNo ratings yet
- Guyton Vias SensitivasDocument13 pagesGuyton Vias SensitivassaraNo ratings yet
- Reflejos somático-somáticos y su importancia clínicaDocument4 pagesReflejos somático-somáticos y su importancia clínicaNombrenombreNo ratings yet
- CAPITULO 12 CUESTIONARIO SALADIN TEJIDO NERVIOSO Páginas 440Document9 pagesCAPITULO 12 CUESTIONARIO SALADIN TEJIDO NERVIOSO Páginas 440David MontesinosNo ratings yet
- Comisura TelencefalicaDocument6 pagesComisura TelencefalicaHector SuarezNo ratings yet
- Tracto corticoespinal: trayecto y funcionesDocument4 pagesTracto corticoespinal: trayecto y funcionesAle100% (1)
- ANATOMÍA DEL SISTEMA NERVIOSO-preguntasDocument5 pagesANATOMÍA DEL SISTEMA NERVIOSO-preguntas2102110021100% (1)
- La organización cortical somatosensorial: de SI a SIIDocument19 pagesLa organización cortical somatosensorial: de SI a SIIPedro FlorNo ratings yet
- Sistema auditivo: apuntes de neurofisiologíaDocument9 pagesSistema auditivo: apuntes de neurofisiologíaJuan Pablo MoyaNo ratings yet
- Sistema nervioso central y periféricoDocument6 pagesSistema nervioso central y periféricoJavier Felipe NovaNo ratings yet
- Anatomia y Fisiologia Del SNCDocument7 pagesAnatomia y Fisiologia Del SNCRafael Cueva AmayaNo ratings yet
- Cuestionario Del Sistema NerviosoDocument11 pagesCuestionario Del Sistema Nerviosovielmiz95No ratings yet
- Formacion ReticularDocument15 pagesFormacion ReticularitzelNo ratings yet
- Trabajo Pares CranealesDocument54 pagesTrabajo Pares CranealesLuulaa Minguin GarciaNo ratings yet
- Cuestionario III.1 - CavidadesDocument19 pagesCuestionario III.1 - CavidadesPelea Con Pendejos TresmilNo ratings yet
- Sistema nervioso: repaso de funciones y estructurasDocument9 pagesSistema nervioso: repaso de funciones y estructurasmonse barbaNo ratings yet
- Corteza CerebralDocument35 pagesCorteza CerebralGiovanna Uceda60% (5)
- Las NeuronasDocument4 pagesLas NeuronasAnonymous XdwJNaNo ratings yet
- Relacion Medico PacienteDocument4 pagesRelacion Medico PacienteBere MezaNo ratings yet
- Formación del sistema nervioso y estructuras asociadasDocument6 pagesFormación del sistema nervioso y estructuras asociadasGENICITA100% (1)
- DCP-sintomas-diagnosticoDocument8 pagesDCP-sintomas-diagnosticoAlbaNo ratings yet
- El Flujo Axonico o AxonalDocument4 pagesEl Flujo Axonico o AxonalHiginioNo ratings yet
- PRÁCTICA No. 5 TALLO CEREBRAL Dr. Luis Arturo Juárez I. Marzo 2012Document10 pagesPRÁCTICA No. 5 TALLO CEREBRAL Dr. Luis Arturo Juárez I. Marzo 2012Julio PerezNo ratings yet
- 5-1-Vias de Conduccion ResumenDocument21 pages5-1-Vias de Conduccion ResumenAriadna Wolf71% (7)
- Conducción del sonido desde la membrana timpánica hasta la cócleaDocument6 pagesConducción del sonido desde la membrana timpánica hasta la cócleaSarely FernándezNo ratings yet
- Cuadro Pares CranealesDocument6 pagesCuadro Pares Cranealesdavid realesNo ratings yet
- Mapa Mental Sistema Nervioso CentralDocument5 pagesMapa Mental Sistema Nervioso Centralharold a romeroNo ratings yet
- Talamo HipotalamoDocument37 pagesTalamo HipotalamoMachuca EdinsonNo ratings yet
- Histologia de CortezaDocument16 pagesHistologia de Cortezahomeromoron1100% (1)
- GUIA 04 PhysioEx 2019Document16 pagesGUIA 04 PhysioEx 2019jossNo ratings yet
- Contribuciones Del Cerebelo y Los Ganglios Basales Al Control Motor GlobalDocument5 pagesContribuciones Del Cerebelo y Los Ganglios Basales Al Control Motor Globaleladio recalde100% (2)
- Función motora corteza y tronco encéfaloDocument7 pagesFunción motora corteza y tronco encéfaloMirian GutierrezNo ratings yet
- La Médula EspinalDocument15 pagesLa Médula Espinalrocio garciaNo ratings yet
- Glosario de NeuropsicologíaDocument138 pagesGlosario de NeuropsicologíaJu Gzn100% (1)
- Generalidades de Sistema UrinarioDocument7 pagesGeneralidades de Sistema UrinarioRocioNo ratings yet
- CerebeloDocument21 pagesCerebeloNatalia Aquino100% (1)
- Embriologia Del Sistema UrinarioDocument5 pagesEmbriologia Del Sistema UrinarioAlessia BeckerNo ratings yet
- Grácil Cuneado funciones núcleosDocument10 pagesGrácil Cuneado funciones núcleosAtilio LópezNo ratings yet
- Organizacion Del Sistema Nervioso 1Document25 pagesOrganizacion Del Sistema Nervioso 1Omar VegaNo ratings yet
- Apuntes Sinapsis 1Document10 pagesApuntes Sinapsis 1Vanessa MuñozNo ratings yet
- 01 Diseño General Del SNCDocument18 pages01 Diseño General Del SNCElvis SantayNo ratings yet
- LeucemiasDocument4 pagesLeucemiasrodrigo4590No ratings yet
- Hoja Recomendaciones EDADocument2 pagesHoja Recomendaciones EDArodrigo4590No ratings yet
- Solicitud de Exámenes de LaboratorioDocument2 pagesSolicitud de Exámenes de Laboratoriorodrigo4590No ratings yet
- Análisis de ArtículoDocument8 pagesAnálisis de Artículorodrigo4590No ratings yet
- ChecoDocument18 pagesChecorodrigo4590No ratings yet
- DefinicionesDocument4 pagesDefinicionesrodrigo4590No ratings yet
- Ascitis y EsputoDocument17 pagesAscitis y Esputorodrigo4590No ratings yet
- Trastornos Hematológicos Del Recién NacidoDocument30 pagesTrastornos Hematológicos Del Recién Nacidorodrigo4590100% (1)
- CefaleasDocument6 pagesCefaleasrodrigo4590No ratings yet
- Atención Primaria de La SaludDocument2 pagesAtención Primaria de La Saludrodrigo4590No ratings yet
- Análisis de Artículo PRISMADocument3 pagesAnálisis de Artículo PRISMArodrigo4590No ratings yet
- Cap 13 Sistema EndócrinoDocument10 pagesCap 13 Sistema Endócrinorodrigo4590No ratings yet
- GlosarioDocument1 pageGlosariorodrigo4590No ratings yet
- Análisis de DecisionesDocument16 pagesAnálisis de Decisionesrodrigo4590No ratings yet
- Casos ClínicosDocument7 pagesCasos Clínicosrodrigo4590No ratings yet
- Análisis de SobrevidaDocument1 pageAnálisis de Sobrevidarodrigo4590No ratings yet
- Estadística InferencialDocument16 pagesEstadística Inferencialrodrigo4590No ratings yet
- Cap 6 Tejido ConectivoDocument10 pagesCap 6 Tejido Conectivorodrigo4590100% (1)
- Cap 7 Cartílago y HuesoDocument18 pagesCap 7 Cartílago y Huesorodrigo4590No ratings yet
- Cap 5 Epitelio y GlándulasDocument14 pagesCap 5 Epitelio y Glándulasrodrigo4590No ratings yet
- Cap 46 Receptores Sensitivos, Circuitos Neuronales para El Procesamiento de La InformaciónDocument8 pagesCap 46 Receptores Sensitivos, Circuitos Neuronales para El Procesamiento de La Informaciónrodrigo4590No ratings yet
- Cap 11 Sistema CirculatorioDocument7 pagesCap 11 Sistema Circulatoriorodrigo4590No ratings yet
- Cap 1 Introducción A La Histología y Técnicas Histológicas BásicasDocument8 pagesCap 1 Introducción A La Histología y Técnicas Histológicas Básicasrodrigo4590100% (2)
- Declaración CONSORTDocument2 pagesDeclaración CONSORTrodrigo4590No ratings yet
- Cap 10 Sangre y HematopoyesisDocument12 pagesCap 10 Sangre y Hematopoyesisrodrigo4590No ratings yet
- Cap 3 NúcleoDocument9 pagesCap 3 Núcleorodrigo4590No ratings yet
- Sensibilidad somática: receptores táctiles y vías de transmisiónDocument8 pagesSensibilidad somática: receptores táctiles y vías de transmisiónrodrigo4590100% (1)
- Cap 6 Contracción Del Músculo EsqueléticoDocument9 pagesCap 6 Contracción Del Músculo Esqueléticorodrigo4590No ratings yet
- Cap 4 Matriz ExtracelularDocument5 pagesCap 4 Matriz Extracelularrodrigo4590No ratings yet
- Sistema nervioso, sinapsis y neurotransmisoresDocument11 pagesSistema nervioso, sinapsis y neurotransmisoresrodrigo4590100% (2)
- FICHA DIAGNÓSTICA Arte y Cultura 1ºy 2° SECDocument4 pagesFICHA DIAGNÓSTICA Arte y Cultura 1ºy 2° SECErnesto BermudezNo ratings yet
- Memoria Gemchi 2014 Subd TecDocument132 pagesMemoria Gemchi 2014 Subd TecSergio GalvezNo ratings yet
- Experiencia cercana a la muerte revela el amor de DiosDocument68 pagesExperiencia cercana a la muerte revela el amor de DiosCeci SierraNo ratings yet
- Listado Farmacos InmunosupresoresDocument2 pagesListado Farmacos InmunosupresoresAdolfo PinoNo ratings yet
- Clase 7 Plantas VascularesDocument17 pagesClase 7 Plantas VascularesAna UrbanoNo ratings yet
- Manejo Prehospitalario Del ShockDocument20 pagesManejo Prehospitalario Del ShockDarwin PeñaNo ratings yet
- HOMINIZACIONDocument4 pagesHOMINIZACIONPaula CalderónNo ratings yet
- Ca3 - Pre Comisonado y Comisionamiento - Etapa Ii Relavera B4 Rev1Document13 pagesCa3 - Pre Comisonado y Comisionamiento - Etapa Ii Relavera B4 Rev1richard_manayayNo ratings yet
- ED-5339 EnspDocument11 pagesED-5339 EnspJoel MartinezNo ratings yet
- Dieta alcalina puede mejorar la salud ósea, muscular y renalDocument31 pagesDieta alcalina puede mejorar la salud ósea, muscular y renalAngeles LagosNo ratings yet
- Centrales U5 PDFDocument15 pagesCentrales U5 PDFRaul Montante SalazarNo ratings yet
- Plan de Control de Calidad SHPDocument80 pagesPlan de Control de Calidad SHPMauricio FachinNo ratings yet
- José Luis Alvarado: Servidumbre Humana. William Somerset MaughamDocument3 pagesJosé Luis Alvarado: Servidumbre Humana. William Somerset MaughamDiego Cruz SantiagoNo ratings yet
- Proyecto Bt. Tampico Obdulio Cesar SanchezDocument66 pagesProyecto Bt. Tampico Obdulio Cesar SanchezAbrahamPartidaNo ratings yet
- A8envejecimiento Adulto Mayor y Mujer Climatérica.Document19 pagesA8envejecimiento Adulto Mayor y Mujer Climatérica.Susana HerreraNo ratings yet
- Formacion VirtudesDocument10 pagesFormacion VirtudesOrlando Jose HerediaNo ratings yet
- Impacto AmbientalDocument16 pagesImpacto AmbientalJesus CarbajalNo ratings yet
- Sistema MaterialDocument6 pagesSistema MaterialAlejandra Romero FracalossiNo ratings yet
- Area Clinica - El Tratamiento Del Padecimiento Subjetivo en La Experiencia AnalíticaDocument9 pagesArea Clinica - El Tratamiento Del Padecimiento Subjetivo en La Experiencia Analíticamblanco15No ratings yet
- Pancreatitis en PediatriaDocument15 pagesPancreatitis en PediatriaMabelcita Rivera AndiaNo ratings yet
- Curva de MiscibilidadDocument7 pagesCurva de MiscibilidadAlvarez Reyes YahirNo ratings yet
- Protocolo de Paso de Cateter Central de InsercionDocument14 pagesProtocolo de Paso de Cateter Central de InsercionJENNYNo ratings yet
- Pregunta 2Document2 pagesPregunta 2Edith VNo ratings yet
- Terapia Génica Equipo 5Document15 pagesTerapia Génica Equipo 5Jesse DiazNo ratings yet
- Memoria 2019 PronacaDocument77 pagesMemoria 2019 PronacaAmparo MoreiraNo ratings yet
- Metodologia Del Genoma HumanoDocument5 pagesMetodologia Del Genoma Humanoubi_rNo ratings yet
- Valoración Neurológica 3Document49 pagesValoración Neurológica 3GABRIEL FLORES ARREDONDO100% (1)
- Cuídate para CuidarDocument2 pagesCuídate para CuidarDocor Comunicacion100% (1)
- Factores Que Afectan Al Indice de Productividad.Document6 pagesFactores Que Afectan Al Indice de Productividad.Jannela Tumbaco Aguayo100% (1)