You might also like
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Wolbachia Write Up For IPTKFINAL PDFDocument13 pagesWolbachia Write Up For IPTKFINAL PDFdzc2016No ratings yet
- Wolbachia Strain Reduces Transmission of Zika VirusDocument7 pagesWolbachia Strain Reduces Transmission of Zika Virusdzc2016No ratings yet
- Field Evaluation of The Establishment Wolbachia Project in VietnamDocument14 pagesField Evaluation of The Establishment Wolbachia Project in VietnamNik FajarNo ratings yet
- Wolbachia SpreadDocument2 pagesWolbachia Spreaddzc2016No ratings yet
- What Makes Community Engagement Effective2Document19 pagesWhat Makes Community Engagement Effective2dzc2016No ratings yet
- Community Participation in Dengue Control in BrazilDocument10 pagesCommunity Participation in Dengue Control in Brazildzc2016No ratings yet
- Invasion of Wolbachia at The Residential Block Level Is Associated With Local Abundance of Stegomyia and Property AttributesDocument8 pagesInvasion of Wolbachia at The Residential Block Level Is Associated With Local Abundance of Stegomyia and Property Attributesdzc2016No ratings yet
- Designing A Community Engagement FrameworkDocument12 pagesDesigning A Community Engagement Frameworkdzc2016No ratings yet
- Wolbachia - ZikaDocument5 pagesWolbachia - Zikadzc2016No ratings yet
- Assessing Key Safety Concerns of A Wolbachia-Based Strategy in BrazilDocument8 pagesAssessing Key Safety Concerns of A Wolbachia-Based Strategy in Brazildzc2016No ratings yet
- Broader Prevalence of Wolbachia in Insects Including Potential Human Disease VectorsDocument11 pagesBroader Prevalence of Wolbachia in Insects Including Potential Human Disease Vectorsdzc2016No ratings yet
- Successful Establishment of Wolbachia in Aedes Populations To Surppress Dengue Transmission PDFDocument6 pagesSuccessful Establishment of Wolbachia in Aedes Populations To Surppress Dengue Transmission PDFdzc2016No ratings yet
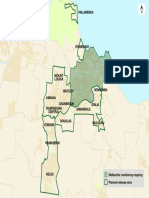
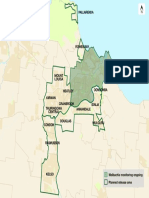
- Townsville Release MapDocument1 pageTownsville Release Mapdzc2016No ratings yet
- Successful Establishment of Wolbachia in Aedes Populations To Surppress Dengue Transmission PDFDocument6 pagesSuccessful Establishment of Wolbachia in Aedes Populations To Surppress Dengue Transmission PDFdzc2016No ratings yet
- Stability of The Wmel Wolbachia Infection Following Invasion Into Aedes Aegypti Population PDFDocument9 pagesStability of The Wmel Wolbachia Infection Following Invasion Into Aedes Aegypti Population PDFdzc2016No ratings yet
- Risk Associated With The Release of Wolbachia-Infected Aedes Aegypti Mosquitoes Into The Environment in An Effort To Control Dengue PDFDocument12 pagesRisk Associated With The Release of Wolbachia-Infected Aedes Aegypti Mosquitoes Into The Environment in An Effort To Control Dengue PDFdzc2016No ratings yet
- Rear and Release A New Paradigm For Dengue ControlDocument5 pagesRear and Release A New Paradigm For Dengue Controldzc2016No ratings yet
- Risk Associated With The Release of Wolbachia-Infected Aedes Aegypti Mosquitoes Into The Environment in An Effort To Control Dengue PDFDocument12 pagesRisk Associated With The Release of Wolbachia-Infected Aedes Aegypti Mosquitoes Into The Environment in An Effort To Control Dengue PDFdzc2016No ratings yet
- Successful Establishment of Wolbachia in Aedes Populations To Surppress Dengue TransmissionDocument1 pageSuccessful Establishment of Wolbachia in Aedes Populations To Surppress Dengue Transmissiondzc2016No ratings yet
- Wolbachia Write Up For IPTKFINAL PDFDocument13 pagesWolbachia Write Up For IPTKFINAL PDFdzc2016No ratings yet
- Stability of The Wmel Wolbachia Infection Following Invasion Into Aedes Aegypti Population PDFDocument9 pagesStability of The Wmel Wolbachia Infection Following Invasion Into Aedes Aegypti Population PDFdzc2016No ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Agarwal - Manual of Neuro OphthalmogyDocument272 pagesAgarwal - Manual of Neuro Ophthalmogythycoon100% (4)
- Jurnal Bahasa InggrisDocument9 pagesJurnal Bahasa InggrisNur Riza Maulidina100% (1)
- Roadmap For TelemedicineDocument48 pagesRoadmap For TelemedicinePradip Gupta100% (2)
- Environmental Studies-FIRST UNIT-vkmDocument59 pagesEnvironmental Studies-FIRST UNIT-vkmRandomNo ratings yet
- Celecoxib, Etoricoxib, DiclofenacDocument13 pagesCelecoxib, Etoricoxib, Diclofenacbaitur rohmahNo ratings yet
- A Comparative Clinical Evaluation of Eranda Taila Matra Basti With Combination of Mefenamic Acid and Dicyclomine in The Management of Udavartini W.S.R To Primary DysmenorrheaDocument8 pagesA Comparative Clinical Evaluation of Eranda Taila Matra Basti With Combination of Mefenamic Acid and Dicyclomine in The Management of Udavartini W.S.R To Primary DysmenorrheaEditor IJTSRDNo ratings yet
- Virtual Care: Advancing The Practice of Veterinary Medicine: Advances in Small Animal Medicine and Surgery April 2019Document4 pagesVirtual Care: Advancing The Practice of Veterinary Medicine: Advances in Small Animal Medicine and Surgery April 2019Anonymous TDI8qdYNo ratings yet
- Mapeh: Music - Arts - Physical Education - HealthDocument23 pagesMapeh: Music - Arts - Physical Education - HealthExtremelydarkness100% (1)
- Plural of Medical Terms UsmpDocument2 pagesPlural of Medical Terms UsmpUSMP FN ARCHIVOSNo ratings yet
- ResearchDocument28 pagesResearchReylon PachesNo ratings yet
- Inheritance Biology EDUNCLEDocument54 pagesInheritance Biology EDUNCLEEvita Almeida100% (1)
- 13) Technical Guideline On Irritation, Sensitization and Hemolysis Study For Chemical DrugsDocument36 pages13) Technical Guideline On Irritation, Sensitization and Hemolysis Study For Chemical DrugsAzam DanishNo ratings yet
- Parathyroi D HormoneDocument25 pagesParathyroi D HormoneChatie PipitNo ratings yet
- Full Text of RA 7719Document4 pagesFull Text of RA 7719camz_hernandezNo ratings yet
- Tap Presentation - Google SlidesDocument12 pagesTap Presentation - Google Slidesapi-208317298No ratings yet
- Bahasa InggrisDocument6 pagesBahasa InggrisSofi SusantoNo ratings yet
- Dr. Sudarto Sppd's FileDocument22 pagesDr. Sudarto Sppd's FileVindy CesarianaNo ratings yet
- Sri Lanka - Averting A National Nutrition Anomaly 2Document16 pagesSri Lanka - Averting A National Nutrition Anomaly 2Rushan LakdimuthuNo ratings yet
- Sample Questionnaire For Understanding The Self: For Private Use OnlyDocument6 pagesSample Questionnaire For Understanding The Self: For Private Use Onlyirene ilustrisimo100% (1)
- 10a General Pest Control Study GuideDocument128 pages10a General Pest Control Study GuideRomeo Baoson100% (1)
- ETT IntubationDocument27 pagesETT IntubationGilbert Sterling Octavius100% (1)
- Hempseed As A Nutritional Resource: An Overview: J.C. CallawayDocument8 pagesHempseed As A Nutritional Resource: An Overview: J.C. CallawayCarmen Yuliana GutierrezNo ratings yet
- Escala de Apatia de StarksteinDocument6 pagesEscala de Apatia de StarksteinVanessa HernandezNo ratings yet
- The Only Book A Teen Will EVER Need To Lose Fat & Build Muscle in The GymDocument146 pagesThe Only Book A Teen Will EVER Need To Lose Fat & Build Muscle in The GymJake Farrugia100% (1)
- Test Bank For Microbiology Basic and Clinical Principles 1st Edition Lourdes P Norman MckayDocument36 pagesTest Bank For Microbiology Basic and Clinical Principles 1st Edition Lourdes P Norman Mckayshriekacericg31u3100% (43)
- Systema Let Every BreathDocument82 pagesSystema Let Every BreathB Agel100% (1)
- Malawi Clinical HIV Guidelines 2019 Addendumversion 8.1Document28 pagesMalawi Clinical HIV Guidelines 2019 Addendumversion 8.1INNOCENT KHULIWANo ratings yet
- GENERAL RISK ASSESSMENT Mechatronics LaboratoryDocument2 pagesGENERAL RISK ASSESSMENT Mechatronics LaboratoryJason TravisNo ratings yet
- Polysaccaride Krestin (PSK)Document5 pagesPolysaccaride Krestin (PSK)Andromeda AbrahamNo ratings yet
- A Manual 20199 CompleetDocument201 pagesA Manual 20199 CompleetJanPerezNo ratings yet