You might also like
- American Fern SocietyDocument14 pagesAmerican Fern Societykhoa128No ratings yet
- Books: Handbook of Storage Tank Systems: Codes, Regulations, and Designs, 1st EditionDocument4 pagesBooks: Handbook of Storage Tank Systems: Codes, Regulations, and Designs, 1st EditionGabriella WidjajaNo ratings yet
- 080 HistorylysimeterdesignDocument10 pages080 HistorylysimeterdesignMarcelo Luz MatosNo ratings yet
- A Model For Gas-Liquid Slug Flow in Horizontal and Near Horizontal TubesDocument11 pagesA Model For Gas-Liquid Slug Flow in Horizontal and Near Horizontal TubesEugene CommerellNo ratings yet
- Dendro ReportDocument13 pagesDendro ReportDr Richard HaddleseyNo ratings yet
- Textile Polypropylene Stents as Artery ImplantsDocument20 pagesTextile Polypropylene Stents as Artery ImplantsKanupriya ChaudharyNo ratings yet
- An Innovative Piston Corer For Large-Volume Sediment Samples: Large-Diameter Piston CorerDocument21 pagesAn Innovative Piston Corer For Large-Volume Sediment Samples: Large-Diameter Piston CorerIlham A RaziqNo ratings yet
- Poepping2004 PDFDocument12 pagesPoepping2004 PDFLuis ManterolaNo ratings yet
- Degradation (Or Lack Thereof) and Drag Reduction of HPAM During Transport in PipelinesDocument14 pagesDegradation (Or Lack Thereof) and Drag Reduction of HPAM During Transport in Pipelinesfabian diazNo ratings yet
- A Comparative Study of Vascular Injection FluidsDocument9 pagesA Comparative Study of Vascular Injection FluidsscribdarnabNo ratings yet
- Evaluation of Fatigue Models - Nigerian StandardDocument12 pagesEvaluation of Fatigue Models - Nigerian StandardDATEME ABAMNo ratings yet
- SPE-182643-MS Pore Size Variability and Sieving Effect in Liquid Shale-A Multiple Permeability Approach and Eagle Ford Case StudyDocument25 pagesSPE-182643-MS Pore Size Variability and Sieving Effect in Liquid Shale-A Multiple Permeability Approach and Eagle Ford Case StudyZhouNo ratings yet
- Chien 1983 Aquacultural-EngineeringDocument3 pagesChien 1983 Aquacultural-EngineeringJorge RodriguezNo ratings yet
- Effects of Stemming Size Distribution On Explosive - C J KonyaDocument4 pagesEffects of Stemming Size Distribution On Explosive - C J KonyarakoNo ratings yet
- Complementario 1Document14 pagesComplementario 1Alondra CodoceoNo ratings yet
- SCR Oddie ShiDocument32 pagesSCR Oddie ShiFrancisco OppsNo ratings yet
- ThomsMegalithicYard 1977Document9 pagesThomsMegalithicYard 1977JNo ratings yet
- Effect of Spreader and Accessory Cone Size On Density of Obturation Using Conventional or Mechanical Lateral CondensationDocument4 pagesEffect of Spreader and Accessory Cone Size On Density of Obturation Using Conventional or Mechanical Lateral CondensationMohammed LafiNo ratings yet
- Influence of Groyne Permeability on Sediment TrappingDocument15 pagesInfluence of Groyne Permeability on Sediment TrappingAddel Yupanqui MirandaNo ratings yet
- 174 Zukoski EE 1966Document17 pages174 Zukoski EE 1966Darryan DhanpatNo ratings yet
- Aspx PDFDocument120 pagesAspx PDFAsem AhmedNo ratings yet
- IADC/SPE 133735 Optimizing in Four Steps Composite Lost-Circulation Pills Without Knowing Loss Zone WidthDocument13 pagesIADC/SPE 133735 Optimizing in Four Steps Composite Lost-Circulation Pills Without Knowing Loss Zone WidthmiladNo ratings yet
- Element variations in rhyolitic magma from gas transportDocument16 pagesElement variations in rhyolitic magma from gas transportclarklipmanNo ratings yet
- Endodontic SealersDocument146 pagesEndodontic SealersSoham DattaNo ratings yet
- RBC ReptilesDocument6 pagesRBC ReptilesnejraNo ratings yet
- SPE 63078 Using The Cased-Hole Formation Tester Tool For Pressure Transient AnalysisDocument15 pagesSPE 63078 Using The Cased-Hole Formation Tester Tool For Pressure Transient Analysisamramazon88No ratings yet
- FulltextDocument9 pagesFulltextwaloodygalbyNo ratings yet
- A New HydraDocument3 pagesA New Hydrajorge solieseNo ratings yet
- Comparison of Methods For Monitoring Reptiles and Amphibians in Upland Forests of The Ouachita MountainsDocument6 pagesComparison of Methods For Monitoring Reptiles and Amphibians in Upland Forests of The Ouachita MountainsManelich TaviraNo ratings yet
- Horizontal Saddle Supported Storage Vessels: A Parametric Study of Plastic Collapse LoadsDocument27 pagesHorizontal Saddle Supported Storage Vessels: A Parametric Study of Plastic Collapse LoadsBimaNo ratings yet
- Knot Theory: The Effects of Different String Sizes On The Probability That Complex Knots Will FormDocument1 pageKnot Theory: The Effects of Different String Sizes On The Probability That Complex Knots Will FormuhfsteNo ratings yet
- Torneck Part IDocument9 pagesTorneck Part IEfren FloresNo ratings yet
- BioRes 06 1 0672 Laitinen KSN Frac Pulp Paper Particles Size 1341Document14 pagesBioRes 06 1 0672 Laitinen KSN Frac Pulp Paper Particles Size 1341jagadish subburajNo ratings yet
- A Comparison of Three Bracket Bases: An in Vitro Study: British Journal of OrthodonticsDocument8 pagesA Comparison of Three Bracket Bases: An in Vitro Study: British Journal of Orthodonticssajida khanNo ratings yet
- 28 Arzani Idesia 31 1Document8 pages28 Arzani Idesia 31 1Septian RizkyNo ratings yet
- Arthroscpic InstrumentsDocument8 pagesArthroscpic InstrumentsRigid EndoscopyNo ratings yet
- The University of Chicago Press International Journal of Plant SciencesDocument8 pagesThe University of Chicago Press International Journal of Plant SciencesGilberto Aleman SancheschulzNo ratings yet
- 5.wonderful World of Endodontic Working Width The Forgotten Dimension - A ReviewDocument7 pages5.wonderful World of Endodontic Working Width The Forgotten Dimension - A ReviewSiva Kumar100% (1)
- Schlesinger 1977Document17 pagesSchlesinger 1977Nathania PalsuNo ratings yet
- Paleostress Inversion Techniques: Methods and Applications for TectonicsFrom EverandPaleostress Inversion Techniques: Methods and Applications for TectonicsNo ratings yet
- Kessler 1983Document9 pagesKessler 1983mostkholy8267No ratings yet
- 1433 FullDocument15 pages1433 FullcubanosNo ratings yet
- Winchester Et Al., 2003 - Dendrogeomorphological ApplicationsDocument21 pagesWinchester Et Al., 2003 - Dendrogeomorphological ApplicationspchiroiuNo ratings yet
- Gill Morphometrics and Gas ExchangeDocument11 pagesGill Morphometrics and Gas Exchangewindy patadunganNo ratings yet
- Dry and Wet Weight of Wheat StrawDocument8 pagesDry and Wet Weight of Wheat StrawYasir NasimNo ratings yet
- 2010 58 4 Emiroglu 245Document16 pages2010 58 4 Emiroglu 245Raju ShresthaNo ratings yet
- Fluid Induced Vibration of Composite Natural Gas PipelinesDocument16 pagesFluid Induced Vibration of Composite Natural Gas PipelinesSimon Achink LubisNo ratings yet
- Research On Drainage MaterialsDocument90 pagesResearch On Drainage MaterialsEFREN BACOLOD MONTALES JRNo ratings yet
- Three Methods of Obturation Using The Obtura II SystemDocument3 pagesThree Methods of Obturation Using The Obtura II SystemAnn M.No ratings yet
- Modelling Droplet Breakage in an Oscillatory Baed ReactorDocument12 pagesModelling Droplet Breakage in an Oscillatory Baed ReactorjohanpenuelaNo ratings yet
- Influence of Vane Size and Equipment On The Results in The Vane Shear TestDocument7 pagesInfluence of Vane Size and Equipment On The Results in The Vane Shear TestNabucodonosor1980No ratings yet
- SPE-170698-MS Rate Transient and Decline Curve Analyses For Continuously (Dual-Porosity) and Discretely Naturally Fractured ReservoirsDocument24 pagesSPE-170698-MS Rate Transient and Decline Curve Analyses For Continuously (Dual-Porosity) and Discretely Naturally Fractured Reservoirsdorian.axel.pt100% (1)
- Visweswar Bhattacharya, Rajesh Kumar WattsDocument6 pagesVisweswar Bhattacharya, Rajesh Kumar WattsViswanath_Poli_2693No ratings yet
- Andrew D. MiallDocument48 pagesAndrew D. MiallAjesh ViswanathanNo ratings yet
- Kristof HanecaDocument16 pagesKristof HanecaBalázs DócziNo ratings yet
- TC TheColdCorrugatorDocument5 pagesTC TheColdCorrugatortikkecoNo ratings yet
- A New Subspecies of the Black Myotis (Bat) from Eastern MexicoFrom EverandA New Subspecies of the Black Myotis (Bat) from Eastern MexicoNo ratings yet
- A New Long-eared Myotis (Myotis Evotis) From Northeastern MexicoFrom EverandA New Long-eared Myotis (Myotis Evotis) From Northeastern MexicoNo ratings yet
- A Procedure For Standardizing Comparative Leaf AnaDocument32 pagesA Procedure For Standardizing Comparative Leaf AnaGilberto Aleman SancheschulzNo ratings yet
- Hydrogel Control of Xylem Hydraulic Resistance in PlantsDocument4 pagesHydrogel Control of Xylem Hydraulic Resistance in PlantsGilberto Aleman SancheschulzNo ratings yet
- Hydraulics of Asteroxylon Mackei An Earl PDFDocument10 pagesHydraulics of Asteroxylon Mackei An Earl PDFGilberto Aleman SancheschulzNo ratings yet
- The University of Chicago Press International Journal of Plant SciencesDocument8 pagesThe University of Chicago Press International Journal of Plant SciencesGilberto Aleman SancheschulzNo ratings yet
- T H I S V I Ew o F L I FeDocument1 pageT H I S V I Ew o F L I FeGilberto Aleman SancheschulzNo ratings yet
- Jansen Et Al-2015-New PhytologistDocument4 pagesJansen Et Al-2015-New PhytologistGilberto Aleman SancheschulzNo ratings yet
- Mercury Uptake and Effects On Growth in Jatropha CurcasDocument6 pagesMercury Uptake and Effects On Growth in Jatropha CurcasGilberto Aleman SancheschulzNo ratings yet
- Vessels in The GramineaeDocument9 pagesVessels in The GramineaeGilberto Aleman SancheschulzNo ratings yet
- Histochemistry General and SpecialDocument56 pagesHistochemistry General and SpecialGilberto Aleman SancheschulzNo ratings yet
- Dlxon's Transpiration and The Ascent of Sap in PlantsDocument4 pagesDlxon's Transpiration and The Ascent of Sap in PlantsGilberto Aleman SancheschulzNo ratings yet
- Ewers & Zimmermann-1984aDocument7 pagesEwers & Zimmermann-1984aGilberto Aleman SancheschulzNo ratings yet
- Anatomical Patterns of Aerenchyma in Aquatic Plants 2008Document12 pagesAnatomical Patterns of Aerenchyma in Aquatic Plants 2008Gilberto Aleman SancheschulzNo ratings yet
- Mills Modern Business Penmanship PDFDocument79 pagesMills Modern Business Penmanship PDFGilberto Aleman SancheschulzNo ratings yet
- Comparative Performance of Trace Element Bioaccumulation Phragmites Typha and ArundoDocument7 pagesComparative Performance of Trace Element Bioaccumulation Phragmites Typha and ArundoGilberto Aleman SancheschulzNo ratings yet
- Gleason Et Al-2015-New PhytologistDocument14 pagesGleason Et Al-2015-New PhytologistGilberto Aleman SancheschulzNo ratings yet
- Cavitation Resistance in Seedless Vascular Plants - The Structure and Function of Interconduit Pit MembranesDocument10 pagesCavitation Resistance in Seedless Vascular Plants - The Structure and Function of Interconduit Pit MembranesGilberto Aleman SancheschulzNo ratings yet
- Plant Physiology PDFDocument23 pagesPlant Physiology PDFGilberto Aleman SancheschulzNo ratings yet
- Duursma FitplcDocument15 pagesDuursma FitplcGilberto Aleman SancheschulzNo ratings yet
- Ellis 1Document46 pagesEllis 1Gilberto Aleman SancheschulzNo ratings yet
- 4154 - Comparative Foliar Epidermal Studies of Twelve Species in The Family Cyperaceae PDFDocument14 pages4154 - Comparative Foliar Epidermal Studies of Twelve Species in The Family Cyperaceae PDFGilberto Aleman SancheschulzNo ratings yet
- Inter - and Intraspecific Structural Variations Pit Membranes Sano 2005Document8 pagesInter - and Intraspecific Structural Variations Pit Membranes Sano 2005Gilberto Aleman SancheschulzNo ratings yet
- An Introduction To R: W. N. Venables, D. M. Smith and The R Core TeamDocument105 pagesAn Introduction To R: W. N. Venables, D. M. Smith and The R Core TeamMahadiNo ratings yet
- Wood Anatomy Conifer Vs Eudicot WoodsprinterfriendlyDocument55 pagesWood Anatomy Conifer Vs Eudicot WoodsprinterfriendlyGilberto Aleman SancheschulzNo ratings yet
- 1 s2.0 S0160412003002344 Main PDFDocument16 pages1 s2.0 S0160412003002344 Main PDFGilberto Aleman SancheschulzNo ratings yet
- 1 s2.0 S1433831911000722 MainDocument12 pages1 s2.0 S1433831911000722 MainGilberto Aleman SancheschulzNo ratings yet
- 1 s2.0 S0034666712000644 MainDocument23 pages1 s2.0 S0034666712000644 MainGilberto Aleman SancheschulzNo ratings yet
- Development of Constructed WetlandsDocument16 pagesDevelopment of Constructed WetlandsGilberto Aleman SancheschulzNo ratings yet
- Lightsheetz1 Sample-Preparation ZeissDocument34 pagesLightsheetz1 Sample-Preparation ZeissGilberto Aleman SancheschulzNo ratings yet
- Ubiquiti Af60-Xr DatasheetDocument3 pagesUbiquiti Af60-Xr Datasheetayman rifaiNo ratings yet
- Comparing Financial Performance of Conventional and Islamic BanksDocument9 pagesComparing Financial Performance of Conventional and Islamic BanksIkbal HardiyantoNo ratings yet
- Silicone Bonding BrochureDocument4 pagesSilicone Bonding BrochureAmir ShahzadNo ratings yet
- Soft StarterDocument6 pagesSoft StarterEric Maglinte TolosaNo ratings yet
- Rethinking Classification and Localization For Object DetectionDocument13 pagesRethinking Classification and Localization For Object DetectionShah Nawaz KhanNo ratings yet
- Remembrance 23 Names PDFDocument1 pageRemembrance 23 Names PDFJennifer ThuncherNo ratings yet
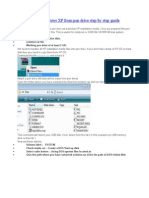
- How To Install Windows XP From Pen Drive Step by Step GuideDocument3 pagesHow To Install Windows XP From Pen Drive Step by Step GuideJithendra Kumar MNo ratings yet
- Project Planning and Management Unit 1Document13 pagesProject Planning and Management Unit 1Savant100% (1)
- Jeremy Hughes ReviewDocument5 pagesJeremy Hughes ReviewgracecavNo ratings yet
- Strata KT Office OpportunityDocument41 pagesStrata KT Office OpportunitySanskar SurekaNo ratings yet
- ANAPHYDocument23 pagesANAPHYYu, Denise Kyla BernadetteNo ratings yet
- EDIBLE VACCINES: A COST-EFFECTIVE SOLUTIONDocument21 pagesEDIBLE VACCINES: A COST-EFFECTIVE SOLUTIONPritish SareenNo ratings yet
- Barry Wylant: Design Thinking and The Experience of InnovationDocument13 pagesBarry Wylant: Design Thinking and The Experience of InnovationVanesa JuarezNo ratings yet
- List of PEGA Interview Questions and AnswersDocument33 pagesList of PEGA Interview Questions and Answersknagender100% (1)
- Security Testing MatDocument9 pagesSecurity Testing MatLias JassiNo ratings yet
- TEI - of - Microsoft - Business Central - FINALDocument23 pagesTEI - of - Microsoft - Business Central - FINALMarketing ComesaNo ratings yet
- LM385Document14 pagesLM385vandocardosoNo ratings yet
- Powers of Central Govt under Environment Protection ActDocument13 pagesPowers of Central Govt under Environment Protection Actsirajudeen INo ratings yet
- Disappearance of Madeleine McCannDocument36 pagesDisappearance of Madeleine McCannCopernicNo ratings yet
- Teaching Support Untuk Managemen HRDocument102 pagesTeaching Support Untuk Managemen HRFernando FmchpNo ratings yet
- Ce125-2500 Open FrameDocument48 pagesCe125-2500 Open FrameRomão OliveiraNo ratings yet
- Primary 2 (Grade 2) - GEP Practice: Contest Problems With Full SolutionsDocument24 pagesPrimary 2 (Grade 2) - GEP Practice: Contest Problems With Full Solutionswenxinyu1002100% (1)
- Effect of Dust On The Performance of Wind Turbines PDFDocument12 pagesEffect of Dust On The Performance of Wind Turbines PDFJallal ArramachNo ratings yet
- PHILHIS Executive Summary - EditedDocument7 pagesPHILHIS Executive Summary - EditedMaxy Bariacto100% (1)
- SPSS-TEST Survey QuestionnaireDocument2 pagesSPSS-TEST Survey QuestionnaireAkshay PatelNo ratings yet
- Expt 1 Yarn Formation (Sherley Trash Analyser)Document7 pagesExpt 1 Yarn Formation (Sherley Trash Analyser)Yashdeep Sharma0% (1)
- Symasym BBDocument37 pagesSymasym BBChandraRizkyNo ratings yet
- History of English Prose PDFDocument21 pagesHistory of English Prose PDFMeisyita QothrunnadaNo ratings yet
- Chams 1Document78 pagesChams 1Das RavindraNo ratings yet
- Marketing Budget: Expenses Q1 Q2 Q3 Q4 Totals Budget %Document20 pagesMarketing Budget: Expenses Q1 Q2 Q3 Q4 Totals Budget %Miaow MiaowNo ratings yet