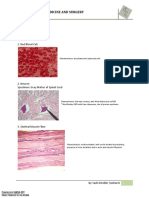
You might also like
- Rapid Identification of Staphylococcus Aureus: FISH Versus PCR MethodsDocument5 pagesRapid Identification of Staphylococcus Aureus: FISH Versus PCR Methodscoleen paraynoNo ratings yet
- Literature Review On Schistosoma HaematobiumDocument7 pagesLiterature Review On Schistosoma Haematobiumfrvkuhrif100% (1)
- Phenotypic CharacterisationDocument7 pagesPhenotypic CharacterisationPraveen KumarNo ratings yet
- Assignment-Statistic Analyzis-Research Method-NugrohoDocument4 pagesAssignment-Statistic Analyzis-Research Method-NugrohoNugroho WiratamaNo ratings yet
- Molecular and biochemical characterization of Staphylococcus aureus β-lactamaseDocument8 pagesMolecular and biochemical characterization of Staphylococcus aureus β-lactamaseMaha hameedNo ratings yet
- Mechanism of Hydrogen Gas Promoted Apoptosis of Lung Adenocarcinoma A549 Cells Through XIAP and BIRC3Document12 pagesMechanism of Hydrogen Gas Promoted Apoptosis of Lung Adenocarcinoma A549 Cells Through XIAP and BIRC3FananiNo ratings yet
- In Vitro Determination of Hydrolytic Enzymes and Echinocandin Susceptibility in Mexican Clinical Isolates of Candida Glabrata Sensu StrictoDocument6 pagesIn Vitro Determination of Hydrolytic Enzymes and Echinocandin Susceptibility in Mexican Clinical Isolates of Candida Glabrata Sensu StrictoFernando EspinosaNo ratings yet
- Biol 230W Lab ReportDocument8 pagesBiol 230W Lab Reportcsh5213No ratings yet
- Reference Intervals For Erythrocyte Sedimentation Rate, Lactate, Fibrinogen, Hematology, and Plasma Protein Electrophoresis in Clinically Healthy Captive Gopher Tortoises (Gopherus Polyphemus)Document9 pagesReference Intervals For Erythrocyte Sedimentation Rate, Lactate, Fibrinogen, Hematology, and Plasma Protein Electrophoresis in Clinically Healthy Captive Gopher Tortoises (Gopherus Polyphemus)Vitor FernandoNo ratings yet
- Genomewide Connection Study Recognizes 2 Brand New Loci Related To AntiNMDAR Encephalitisjpals PDFDocument1 pageGenomewide Connection Study Recognizes 2 Brand New Loci Related To AntiNMDAR Encephalitisjpals PDFcatpail77No ratings yet
- Blood Imp 5Document10 pagesBlood Imp 5ashokNo ratings yet
- 2012 SPN MaldiDocument6 pages2012 SPN MaldiSergey SidorenkoNo ratings yet
- Candida and Candidiasis 2023 Abstract BookDocument209 pagesCandida and Candidiasis 2023 Abstract BooknohemiNo ratings yet
- A Quantitative PCR (Taqman) Assay For Pathogenic: Leptospira SPPDocument22 pagesA Quantitative PCR (Taqman) Assay For Pathogenic: Leptospira SPPVigneshwaran RavishankarNo ratings yet
- Effect of Antibodies On The Expression of Plasmodium Falciparum Circumsporozoite Protein GeneDocument4 pagesEffect of Antibodies On The Expression of Plasmodium Falciparum Circumsporozoite Protein Genereza gomaNo ratings yet
- Alvarez, Et Al. 2013. Developmental and Comparative ImmunologyDocument5 pagesAlvarez, Et Al. 2013. Developmental and Comparative ImmunologyFernandoNo ratings yet
- Citosol (Thiamylal Sodium) Triggers Apoptosis and Affects Gene Expressions of Murine Leukemia RAW 264.7 Cells - RS-C Wu, C-S Yu, 2Document1 pageCitosol (Thiamylal Sodium) Triggers Apoptosis and Affects Gene Expressions of Murine Leukemia RAW 264.7 Cells - RS-C Wu, C-S Yu, 2Alondra MaldonadoNo ratings yet
- Anti-C1q Antibodies: Association With Nephritis and Disease Activity in Systemic Lupus ErythematosusDocument5 pagesAnti-C1q Antibodies: Association With Nephritis and Disease Activity in Systemic Lupus ErythematosusAlisNo ratings yet
- Chlorine Dioxide is a More Effective Disinfectant than Sodium Hypochlorite Against Multi-Drug Resistant BacteriaDocument4 pagesChlorine Dioxide is a More Effective Disinfectant than Sodium Hypochlorite Against Multi-Drug Resistant BacteriaNathalia LedesmaNo ratings yet
- Expression of Fas (CD95:APO-1) and Fas Ligand (FasL) in Experimentally-Induced Acute PancreatitisDocument9 pagesExpression of Fas (CD95:APO-1) and Fas Ligand (FasL) in Experimentally-Induced Acute PancreatitismattpauziNo ratings yet
- Artikel Amdal 2Document18 pagesArtikel Amdal 2Eka PutraNo ratings yet
- Startups That'll Change The Ballya Diagnostic Kit Assembly Machines Industry For The BetterDocument4 pagesStartups That'll Change The Ballya Diagnostic Kit Assembly Machines Industry For The BetterdelodoqpbhNo ratings yet
- BIO 230W Lab ReportDocument8 pagesBIO 230W Lab ReportAlaina WeinheimerNo ratings yet
- 69929oral Most CancersDocument3 pages69929oral Most CancersneriktjpcoNo ratings yet
- Literature Review On BacteriaDocument9 pagesLiterature Review On Bacteriaea83xjp7100% (1)
- Parameters to evaluate the immunostimulant effect of Zymosan A in salmonidsDocument8 pagesParameters to evaluate the immunostimulant effect of Zymosan A in salmonidsThảo ThảoNo ratings yet
- Dissertationen Lmu TiermedizinDocument7 pagesDissertationen Lmu TiermedizinCustomPaperServiceSingapore100% (1)
- Mata 2Document4 pagesMata 2Shofi Nur AidahNo ratings yet
- Literatur Untuk Modul 5 ProfisDocument5 pagesLiteratur Untuk Modul 5 ProfisRisa DedewwNo ratings yet
- Detection de QNR and Aac6 LBCR in Enterobacteriacea IranDocument5 pagesDetection de QNR and Aac6 LBCR in Enterobacteriacea IranclaudiaNo ratings yet
- Protein Extract1Document13 pagesProtein Extract1razor0utNo ratings yet
- Lab Report 3 - Characterization of Color Lake Sediment Soil Bacteria IsolatesDocument13 pagesLab Report 3 - Characterization of Color Lake Sediment Soil Bacteria IsolatesMatthew HeNo ratings yet
- Weil-Felix Test in Diagnosis of ScrubDocument3 pagesWeil-Felix Test in Diagnosis of ScrubvyasakandarpNo ratings yet
- J Med Chem 2004 47 6609-6615-OKDocument7 pagesJ Med Chem 2004 47 6609-6615-OKca_teixeiraNo ratings yet
- Development of Anti-Peptide Polyclonal Antibodies Raised Against Immunogenic Sequences of Protein Encoded by Fj194940.1 GeneDocument9 pagesDevelopment of Anti-Peptide Polyclonal Antibodies Raised Against Immunogenic Sequences of Protein Encoded by Fj194940.1 GeneijsidonlineinfoNo ratings yet
- Differential Diagnosis of Freshwater and Saltwater Drowning Based on Lung Aquaporin-5 ExpressionDocument7 pagesDifferential Diagnosis of Freshwater and Saltwater Drowning Based on Lung Aquaporin-5 ExpressionSGDB10 fightingNo ratings yet
- 2013 Evaluation of The BinaxNOWDocument5 pages2013 Evaluation of The BinaxNOWdatitoxNo ratings yet
- 30 - Evaluation of Prostate-Specific Antigen (PSA) Membrane Test Assays For The Forensic Identification of Seminal Fluid PDFDocument4 pages30 - Evaluation of Prostate-Specific Antigen (PSA) Membrane Test Assays For The Forensic Identification of Seminal Fluid PDFMN IrshadNo ratings yet
- Evaluation Serological Diagnostic Indices For Mucocutaneous Leishmaniasis: Immunofluorescence and Enzyme-Linked For Igm and Iga AntibodiesDocument6 pagesEvaluation Serological Diagnostic Indices For Mucocutaneous Leishmaniasis: Immunofluorescence and Enzyme-Linked For Igm and Iga AntibodiesnisaNo ratings yet
- Food and Chemical Toxicology: Kyoung Jin Nho, Jin Mi Chun, Ho Kyoung KimDocument10 pagesFood and Chemical Toxicology: Kyoung Jin Nho, Jin Mi Chun, Ho Kyoung KimMd Jahidul IslamNo ratings yet
- Anti-Saccharomyces Cerevisiae and Antineutrophil Cytoplasmic Antibodies As Predictors of Inflammatory Bowel DiseaseDocument5 pagesAnti-Saccharomyces Cerevisiae and Antineutrophil Cytoplasmic Antibodies As Predictors of Inflammatory Bowel DiseaseFabio BottariNo ratings yet
- Milestones in Laboratory Procedures and Techniques: Immunohematology BeginningsDocument5 pagesMilestones in Laboratory Procedures and Techniques: Immunohematology BeginningsCatia CorreaNo ratings yet
- Isolation and Analysis of Nephritic Producing Immune Complexes in Plasmodium Berghei Infected MiceDocument7 pagesIsolation and Analysis of Nephritic Producing Immune Complexes in Plasmodium Berghei Infected MicedarknigterNo ratings yet
- Thesis Paper On PneumoniaDocument7 pagesThesis Paper On Pneumoniagbwwdfq5100% (1)
- GRANADOSDocument6 pagesGRANADOSArmando GemeNo ratings yet
- E6CBABB61857Document8 pagesE6CBABB61857RIVALDO JUNINHO ZINEDINE ADIKPETONo ratings yet
- OPA-gag PCRDocument7 pagesOPA-gag PCRmuralidhar_mettaNo ratings yet
- Infections. Molecular Probes For Diagnosis of FungalDocument8 pagesInfections. Molecular Probes For Diagnosis of FungalJes JesNo ratings yet
- Camparison Monovalente AntivenomsDocument1 pageCamparison Monovalente AntivenomsAndrea BaracaldoNo ratings yet
- High Prevalence of Methicillin-Resistant: Staphylococcus Haemolyticus Isolated From Commensals in Healthy AdultsDocument8 pagesHigh Prevalence of Methicillin-Resistant: Staphylococcus Haemolyticus Isolated From Commensals in Healthy AdultsElok NurfaiqohNo ratings yet
- Thesis On Salmonella TyphiDocument4 pagesThesis On Salmonella Typhikimberlyreyessterlingheights100% (2)
- Salivary Tumor MarkersDocument12 pagesSalivary Tumor Markershillary aurenneNo ratings yet
- ALESON - Analysis of The Toxicological and Pharmacokinetic Profile of KaempferolDocument6 pagesALESON - Analysis of The Toxicological and Pharmacokinetic Profile of KaempferolLaísa CordeiroNo ratings yet
- Routine Urine Test May Help Predict Premature Rupture of MembranesDocument10 pagesRoutine Urine Test May Help Predict Premature Rupture of Membranesnadya nazwinnNo ratings yet
- Cold Atmospheric Plasma Discharged in Water and Its Potential Use in Cancer TherapyDocument18 pagesCold Atmospheric Plasma Discharged in Water and Its Potential Use in Cancer TherapySek PyroNo ratings yet
- AsoDocument10 pagesAsoDiah WahyuniNo ratings yet
- Identification of Bacterial Proteins Observed in MALDI TOF Mass Spectra From Whole CellsDocument15 pagesIdentification of Bacterial Proteins Observed in MALDI TOF Mass Spectra From Whole CellsFaisal AliNo ratings yet
- Scientific Proceedings: Detroit, May 21 24, 1946Document22 pagesScientific Proceedings: Detroit, May 21 24, 1946Veneta GizdakovaNo ratings yet
- 69th AACC Annual Scientific Meeting Abstract eBookFrom Everand69th AACC Annual Scientific Meeting Abstract eBookNo ratings yet
- Botany ReviewerDocument23 pagesBotany ReviewerIvy CruzNo ratings yet
- Evolution and Diversity of Woody and Seed Plants: Chapter 5 on LignophytesDocument1 pageEvolution and Diversity of Woody and Seed Plants: Chapter 5 on LignophytesIvy CruzNo ratings yet
- Love, Loss and Life's Deepest QuestionsDocument3 pagesLove, Loss and Life's Deepest QuestionsIvy CruzNo ratings yet
- Muscular System Comparative Anatomy LabDocument2 pagesMuscular System Comparative Anatomy LabIvy CruzNo ratings yet
- 4mm Frog EmbryoDocument3 pages4mm Frog EmbryoIvy CruzNo ratings yet
- Exercise 3 - Embryology LabDocument3 pagesExercise 3 - Embryology LabIvy CruzNo ratings yet
- Exercise 1 - Embryology LabDocument7 pagesExercise 1 - Embryology LabIvy CruzNo ratings yet
- CH 10 MusclesDocument5 pagesCH 10 MusclesIvy CruzNo ratings yet
- Evolution and Diversity of Woody and Seed Plants: Chapter 5 on LignophytesDocument1 pageEvolution and Diversity of Woody and Seed Plants: Chapter 5 on LignophytesIvy CruzNo ratings yet
- Chordates TaxaDocument1 pageChordates TaxaIvy CruzNo ratings yet
- 10mm Frog Embryo - Embryology LabDocument4 pages10mm Frog Embryo - Embryology LabIvy CruzNo ratings yet
- 4mm Frog EmbryoDocument3 pages4mm Frog EmbryoIvy CruzNo ratings yet
- 7mm Frog Embryo - EMBLabDocument3 pages7mm Frog Embryo - EMBLabIvy Cruz100% (1)
- 24 Hour Chick Embryo - Embryology LabDocument3 pages24 Hour Chick Embryo - Embryology LabIvy Cruz50% (2)
- Exercise 2 - Embryology LabDocument2 pagesExercise 2 - Embryology LabIvy CruzNo ratings yet
- Histo 1st ShiftDocument25 pagesHisto 1st ShiftIvy CruzNo ratings yet
- 2017 Year at A Glance PDFDocument1 page2017 Year at A Glance PDFtoanomisNo ratings yet
- Acd 4 Arm and Cubital FossaDocument32 pagesAcd 4 Arm and Cubital FossaIvy CruzNo ratings yet
- CPPI Presentation Borres Et Al.Document19 pagesCPPI Presentation Borres Et Al.Ivy CruzNo ratings yet
- Abdominis Contract To Actively Aid ExhalationDocument4 pagesAbdominis Contract To Actively Aid ExhalationIvy CruzNo ratings yet
- Abdominis Contract To Actively Aid ExhalationDocument4 pagesAbdominis Contract To Actively Aid ExhalationIvy CruzNo ratings yet
- Circulatory Sytem of The CatDocument3 pagesCirculatory Sytem of The CatIvy CruzNo ratings yet
- The Catholic Social Teachings is reputed as the ChurchÕs Best Kept SecretDocument3 pagesThe Catholic Social Teachings is reputed as the ChurchÕs Best Kept SecretIvy CruzNo ratings yet
- Kingdom ProtozoaDocument3 pagesKingdom ProtozoaIvy CruzNo ratings yet
- Mitochondria and Golgi in Adipose CellsDocument3 pagesMitochondria and Golgi in Adipose CellsIvy CruzNo ratings yet
- Gymnosperm TaxaDocument1 pageGymnosperm TaxaIvy CruzNo ratings yet
- Phylum Chordata TaxonomyDocument1 pagePhylum Chordata TaxonomyIvy CruzNo ratings yet
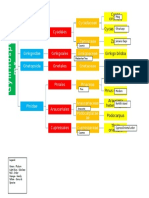
- Legend: Phylum - Violet Class - Blue Subclass - Green Order - Red Family - Orange Genus - Yellow Species - PinkDocument1 pageLegend: Phylum - Violet Class - Blue Subclass - Green Order - Red Family - Orange Genus - Yellow Species - PinkIvy CruzNo ratings yet
- Ana Chem Lab NotesDocument2 pagesAna Chem Lab NotesIvy CruzNo ratings yet
- Bryophytes Taxonomy ReviewerDocument1 pageBryophytes Taxonomy ReviewerIvy CruzNo ratings yet
- Viral Haemorrhagic Fever - Case Investigation Form: Date of Detection of The Case Case IDDocument3 pagesViral Haemorrhagic Fever - Case Investigation Form: Date of Detection of The Case Case IDSalihu MustaphaNo ratings yet
- Voci Assessment OnlineDocument4 pagesVoci Assessment OnlineSaurav DasNo ratings yet
- Final Research Paper 2Document24 pagesFinal Research Paper 2Ahlea PinedaNo ratings yet
- Semis IEMDocument13 pagesSemis IEMVenus Almazan RamiscalNo ratings yet
- Amser Guide To Applying For Radiology Residency: Version 9 - June 2018Document44 pagesAmser Guide To Applying For Radiology Residency: Version 9 - June 2018npr00No ratings yet
- Thesis EgosDocument48 pagesThesis EgosJamie HaravataNo ratings yet
- Malan IDocument24 pagesMalan IJulián Alberto Muñoz FigueroaNo ratings yet
- Quarter 1 - Module 2: Yoga As A Recreational ActivityDocument5 pagesQuarter 1 - Module 2: Yoga As A Recreational ActivityJustine Dave rubaNo ratings yet
- What Is A Warm Up and How To Warm Up ProperlyDocument5 pagesWhat Is A Warm Up and How To Warm Up ProperlyHamilton DurantNo ratings yet
- Ich Guideline q4b Annex 7 r2 Note Evaluation Recommendation Pharmacopoeial Texts Use Ich Regions - enDocument5 pagesIch Guideline q4b Annex 7 r2 Note Evaluation Recommendation Pharmacopoeial Texts Use Ich Regions - enprincess_022No ratings yet
- D-Day: Doug Stebleton Reporting From NormandyDocument12 pagesD-Day: Doug Stebleton Reporting From NormandyBS Central, Inc. "The Buzz"No ratings yet
- The Romantic DiscourseDocument9 pagesThe Romantic Discourse9w6w22b9pmNo ratings yet
- The Idea of Applying The "Broken Windows Theory" As Quality Tool To Improve Risk Management StrategiesDocument8 pagesThe Idea of Applying The "Broken Windows Theory" As Quality Tool To Improve Risk Management StrategiesInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Alphatrak 2 User GuideDocument23 pagesAlphatrak 2 User GuideGlenn MartynNo ratings yet
- The Educational Benefits of Videogames: Mark GriffithsDocument5 pagesThe Educational Benefits of Videogames: Mark GriffithsAngelo ChaustreNo ratings yet
- Case Study For CellulitisDocument21 pagesCase Study For CellulitisMary Ann Lumbay Paye75% (4)
- Mclat Book ReviewDocument22 pagesMclat Book ReviewNezeil Dubduban100% (1)
- Task 2 ExamplesDocument4 pagesTask 2 ExamplescrvenaNo ratings yet
- Paralympics Excellence CentreDocument7 pagesParalympics Excellence CentreNasmal ZubairNo ratings yet
- Macular Buckle AdvancementsDocument4 pagesMacular Buckle AdvancementsMohammad A. BawtagNo ratings yet
- Bai Tap Tieng Anh Lop 8 Unit 6 The Young Pioneers Club So 4Document3 pagesBai Tap Tieng Anh Lop 8 Unit 6 The Young Pioneers Club So 4Nha TranNo ratings yet
- Tagoloan Community College: Outcomes-Based Teaching and Learning Plan in Diploma in MidwiferyDocument2 pagesTagoloan Community College: Outcomes-Based Teaching and Learning Plan in Diploma in MidwiferyRam AugustNo ratings yet
- Mathematics Manual For Water and Wastewater Treatment Plant Operators, Second Edition - Three Volume Set Volume 1 (PDFDrive)Document292 pagesMathematics Manual For Water and Wastewater Treatment Plant Operators, Second Edition - Three Volume Set Volume 1 (PDFDrive)Rianna100% (1)
- Early Warning Alert and Response in Emergencies: An Operational GuideDocument208 pagesEarly Warning Alert and Response in Emergencies: An Operational GuideVitaNo ratings yet
- Art Profe Julian 1Document9 pagesArt Profe Julian 1Julian LoaizaNo ratings yet
- 5550 Wheelhouse Generator ManualDocument16 pages5550 Wheelhouse Generator ManualLuys HernandezNo ratings yet
- Laporan POR 2022 KalibagorDocument15 pagesLaporan POR 2022 Kalibagoribnu an nafisNo ratings yet
- Counselling AssignmentDocument4 pagesCounselling AssignmentMegha BansalNo ratings yet
- Bagong Kalsada Annexes BDTPlan.Document107 pagesBagong Kalsada Annexes BDTPlan.Barangay MabuloNo ratings yet
- Sufism and Mental Health: S. Haque Nizamie Mohammad Zia Ul Haq Katshu N. A. UvaisDocument2 pagesSufism and Mental Health: S. Haque Nizamie Mohammad Zia Ul Haq Katshu N. A. Uvaisblackpetal1No ratings yet