You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- 5 Email Templates v3Document7 pages5 Email Templates v3Kolyo DankovNo ratings yet
- Keith J. Laidler, John H. Meiser - Physical Chemistry-Houghton Mifflin (1999)Document1,050 pagesKeith J. Laidler, John H. Meiser - Physical Chemistry-Houghton Mifflin (1999)chunibyoxdelusions100% (2)
- Use of Surfactants in Metal Cutting FluidsDocument24 pagesUse of Surfactants in Metal Cutting FluidsLen SurbanNo ratings yet
- Universal Law of GravitationDocument13 pagesUniversal Law of GravitationDavid NgNo ratings yet
- Cancer Metabolic Disease Parasites Ketogenic Diet Cancer TherapyDocument3 pagesCancer Metabolic Disease Parasites Ketogenic Diet Cancer TherapyKolyo Dankov100% (2)
- Class XII Physics Support Material Final-1Document399 pagesClass XII Physics Support Material Final-1Adithyan AjithNo ratings yet
- Jiang2015 PDFDocument6 pagesJiang2015 PDFKolyo DankovNo ratings yet
- Direct Observation of Coherent Inter-Orbital Spin-Exchange DynamicsDocument8 pagesDirect Observation of Coherent Inter-Orbital Spin-Exchange DynamicsKolyo DankovNo ratings yet
- Taylor 2015Document7 pagesTaylor 2015Kolyo DankovNo ratings yet
- Goltsev 2009Document16 pagesGoltsev 2009Kolyo DankovNo ratings yet
- Cooperstock 2015Document5 pagesCooperstock 2015Kolyo DankovNo ratings yet
- DRobertsRCurtis Medi1Document15 pagesDRobertsRCurtis Medi1Kolyo DankovNo ratings yet
- Did LIGO Detect Dark Matter?Document6 pagesDid LIGO Detect Dark Matter?Kolyo DankovNo ratings yet
- Local Conformal Symmetry: The Missing Symmetry Component For Space and TimeDocument5 pagesLocal Conformal Symmetry: The Missing Symmetry Component For Space and TimeKolyo DankovNo ratings yet
- Pulsed Electron-Nuclear Double Resonance (Endor) at 140 GHZDocument12 pagesPulsed Electron-Nuclear Double Resonance (Endor) at 140 GHZKolyo DankovNo ratings yet
- Jiang2015 PDFDocument6 pagesJiang2015 PDFKolyo DankovNo ratings yet
- 565920Document12 pages565920Kolyo DankovNo ratings yet
- Erv 272Document10 pagesErv 272Kolyo DankovNo ratings yet
- Pulsed Electron-Nuclear Double Resonance (Endor) at 140 GHZDocument12 pagesPulsed Electron-Nuclear Double Resonance (Endor) at 140 GHZKolyo DankovNo ratings yet
- Viruses: Adenovirus 36 and Obesity: An OverviewDocument22 pagesViruses: Adenovirus 36 and Obesity: An OverviewKolyo DankovNo ratings yet
- 1505 04753v3Document8 pages1505 04753v3Kolyo DankovNo ratings yet
- Soerensen 2016Document25 pagesSoerensen 2016Kolyo DankovNo ratings yet
- Abram Ow Ski 2016Document14 pagesAbram Ow Ski 2016Kolyo DankovNo ratings yet
- ADocument2 pagesAKolyo DankovNo ratings yet
- BIGCHEM AnnouncementDocument1 pageBIGCHEM AnnouncementKolyo DankovNo ratings yet
- ADocument3 pagesAKolyo DankovNo ratings yet
- Watrous - New Pottery From Cave Psychro and Its Implications in Minoan CreteDocument27 pagesWatrous - New Pottery From Cave Psychro and Its Implications in Minoan CreteKolyo DankovNo ratings yet
- Biology of Aging Program-05.9.14Document10 pagesBiology of Aging Program-05.9.14Kolyo DankovNo ratings yet
- KekichDocument9 pagesKekichKolyo DankovNo ratings yet
- Budni 2016Document9 pagesBudni 2016Kolyo DankovNo ratings yet
- Rs If 20130901Document22 pagesRs If 20130901Kolyo DankovNo ratings yet
- Corrosionresistance of Delhi Iron Pillar R Balasubramaniam 2000Document27 pagesCorrosionresistance of Delhi Iron Pillar R Balasubramaniam 2000Srini KalyanaramanNo ratings yet
- Angular Analysis of The B0 Decay UsingDocument79 pagesAngular Analysis of The B0 Decay UsingKolyo DankovNo ratings yet
- Electrochemistry Past Papers 2022-14Document4 pagesElectrochemistry Past Papers 2022-1410 A Pratyush Dubey0% (1)
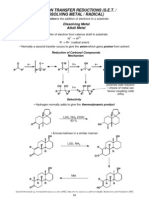
- Metal ReductionDocument11 pagesMetal Reductiondeepthimahanthi sabithaNo ratings yet
- 11 CAPS 11 Student Copy AnanthGarg&On Trak0EduCompetishunDocument8 pages11 CAPS 11 Student Copy AnanthGarg&On Trak0EduCompetishunSepulcher ArmsNo ratings yet
- Chinese Cable Details DataDocument131 pagesChinese Cable Details DataJoyanta DebNo ratings yet
- Sunanda SharmaDocument11 pagesSunanda SharmaPrashant A UNo ratings yet
- Experiment 9 Measurement of Hydraulic Conductivity of Granular SoilsDocument12 pagesExperiment 9 Measurement of Hydraulic Conductivity of Granular SoilsAllyssa SuyatNo ratings yet
- 5054 Space Physics Practice QuestionsDocument14 pages5054 Space Physics Practice QuestionsSoya BeansNo ratings yet
- Metco 54NS-1 (Aluminum Seal Coat) PDFDocument3 pagesMetco 54NS-1 (Aluminum Seal Coat) PDFJ. BangjakNo ratings yet
- US8178353 - Method For Determination of Polymer Concentration in Water SystemsDocument15 pagesUS8178353 - Method For Determination of Polymer Concentration in Water SystemsKhanh Nguyen TuanNo ratings yet
- Earth Science ActivitiesDocument10 pagesEarth Science ActivitiescykenNo ratings yet
- Energy Relationships in Chemical ReactionsDocument11 pagesEnergy Relationships in Chemical ReactionsDanise NicoleNo ratings yet
- CamSur National HS Earth Science Modules 6-8Document3 pagesCamSur National HS Earth Science Modules 6-8Jerald De La CruzNo ratings yet
- Astm D512 - 12 - Cloruros en AguaDocument9 pagesAstm D512 - 12 - Cloruros en AguaEliasNo ratings yet
- Chapter 14Document8 pagesChapter 14nelaojNo ratings yet
- ASTM A295-98 (Obsolete)Document2 pagesASTM A295-98 (Obsolete)techietaddyNo ratings yet
- CMGPS User ManualDocument46 pagesCMGPS User ManualULTG KOTABUMINo ratings yet
- CBSE Class 9 Biology Natural Resources Notes PDFDocument10 pagesCBSE Class 9 Biology Natural Resources Notes PDFAbhishekNo ratings yet
- Answers: Name Mentor Group 2 0 3 0 DateDocument7 pagesAnswers: Name Mentor Group 2 0 3 0 DateBuloi PoijNo ratings yet
- Chemical Bath Deposition: Mark - Deguire@case - EduDocument21 pagesChemical Bath Deposition: Mark - Deguire@case - EduH Cuarto PeñaNo ratings yet
- Most and Least Learned Competencies: Republic of The PhilippinesDocument5 pagesMost and Least Learned Competencies: Republic of The PhilippinesLorena De BelenNo ratings yet
- (G. - P. - Motulevich) Optical Properties of Metals and Intermolecular InteractionsDocument234 pages(G. - P. - Motulevich) Optical Properties of Metals and Intermolecular Interactions张尚宇No ratings yet
- What Is Matter???: Pure Substance MixtureDocument4 pagesWhat Is Matter???: Pure Substance MixtureArdelynNo ratings yet
- 940 U S Face Permeability Monitoring For Gibe III RCC Dam and Resin Injection Works, Pittalis, Cagiano, Pietrangeli, BianDocument8 pages940 U S Face Permeability Monitoring For Gibe III RCC Dam and Resin Injection Works, Pittalis, Cagiano, Pietrangeli, BianDataniel RosarioNo ratings yet
- 2000 - Fukai - Thermal Conductivity Enhancement of Energy Storage Media Using Carbon FibersDocument14 pages2000 - Fukai - Thermal Conductivity Enhancement of Energy Storage Media Using Carbon FibersShiva Kumar NidanampallyNo ratings yet
- Pecora A-103 DataSheet 4-13Document2 pagesPecora A-103 DataSheet 4-13Andres RodriguezNo ratings yet
- 123Document20 pages123Dacy ChowNo ratings yet