You might also like
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Phytotherapy On CancerDocument21 pagesPhytotherapy On CancerSiddhendu Bhattacharjee100% (1)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Apron CapacityDocument10 pagesApron CapacityMuchammad Ulil AidiNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Tutorial 5 SolvedDocument3 pagesTutorial 5 SolvedAshutoshKumarNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Core ValuesDocument1 pageCore ValuesIan Abel AntiverosNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Continue Practice Exam Test Questions Part 1 of The SeriesDocument7 pagesContinue Practice Exam Test Questions Part 1 of The SeriesKenn Earl Bringino VillanuevaNo ratings yet
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Catedral de SevillaDocument28 pagesCatedral de SevillaAnonymous LkgNmKeNo ratings yet
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Skills Checklist - Gastrostomy Tube FeedingDocument2 pagesSkills Checklist - Gastrostomy Tube Feedingpunam todkar100% (1)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Final Project Strategic ManagementDocument2 pagesFinal Project Strategic ManagementMahrukh RasheedNo ratings yet
- Delonghi Esam Series Service Info ItalyDocument10 pagesDelonghi Esam Series Service Info ItalyBrko BrkoskiNo ratings yet
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Catify To Satisfy - Simple Solutions For Creating A Cat-Friendly Home (PDFDrive)Document315 pagesCatify To Satisfy - Simple Solutions For Creating A Cat-Friendly Home (PDFDrive)Paz Libros100% (2)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Winter CrocFest 2017 at St. Augustine Alligator Farm - Final ReportDocument6 pagesWinter CrocFest 2017 at St. Augustine Alligator Farm - Final ReportColette AdamsNo ratings yet
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Mosfet Irfz44Document8 pagesMosfet Irfz44huynhsang1979No ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Cummin C1100 Fuel System Flow DiagramDocument8 pagesCummin C1100 Fuel System Flow DiagramDaniel KrismantoroNo ratings yet
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Apple Change ManagementDocument31 pagesApple Change ManagementimuffysNo ratings yet
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Hidrl1 PDFDocument7 pagesHidrl1 PDFRajesh Kumar100% (1)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- PresentationDocument27 pagesPresentationMenuka WatankachhiNo ratings yet
- English Homework 10 Grammar Focus 2: Lecturer: Mr. Dr. H. Abdul Hamid, M.SiDocument4 pagesEnglish Homework 10 Grammar Focus 2: Lecturer: Mr. Dr. H. Abdul Hamid, M.SiMutiara siwa UtamiNo ratings yet
- Comparison Between CompetitorsDocument2 pagesComparison Between Competitorsritesh singhNo ratings yet
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
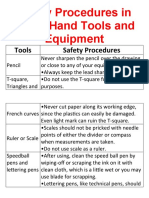
- Safety Procedures in Using Hand Tools and EquipmentDocument12 pagesSafety Procedures in Using Hand Tools and EquipmentJan IcejimenezNo ratings yet
- KsDocument5 pagesKsnurlatifahNo ratings yet
- Julia Dito ResumeDocument3 pagesJulia Dito Resumeapi-253713289No ratings yet
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Guyana and The Islamic WorldDocument21 pagesGuyana and The Islamic WorldshuaibahmadkhanNo ratings yet
- Peer PressureDocument13 pagesPeer PressuremightymarcNo ratings yet
- Neonatal Mortality - A Community ApproachDocument13 pagesNeonatal Mortality - A Community ApproachJalam Singh RathoreNo ratings yet
- Rajiv Gandhi University of Health Sciences, Bengaluru, KarnatakaDocument9 pagesRajiv Gandhi University of Health Sciences, Bengaluru, KarnatakaNavin ChandarNo ratings yet
- PFEIFER Angled Loops For Hollow Core Slabs: Item-No. 05.023Document1 pagePFEIFER Angled Loops For Hollow Core Slabs: Item-No. 05.023adyhugoNo ratings yet
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Rifle May 2015 USADocument72 pagesRifle May 2015 USAhanshcNo ratings yet
- Fundaciones Con PilotesDocument48 pagesFundaciones Con PilotesReddy M.Ch.No ratings yet
- Evs ProjectDocument19 pagesEvs ProjectSaloni KariyaNo ratings yet
- Pg2022 ResultDocument86 pagesPg2022 ResultkapilNo ratings yet
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)