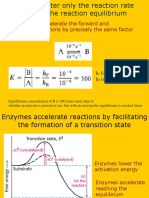
You might also like
- Mechanism of Enzyme ActionDocument2 pagesMechanism of Enzyme ActionBlazy InhumangNo ratings yet
- Enzymes: Properties and KineticsDocument59 pagesEnzymes: Properties and Kineticsthamizh555100% (2)
- Quantitative Human Physiology: An IntroductionFrom EverandQuantitative Human Physiology: An IntroductionRating: 2 out of 5 stars2/5 (1)
- Mechanism of Enzyme CatalysisDocument18 pagesMechanism of Enzyme CatalysisBuhlebenkosi Jackie BhebheNo ratings yet
- Allosteric Enzymes and Feedback InhibitionDocument37 pagesAllosteric Enzymes and Feedback InhibitionLiana ChowdhuryNo ratings yet
- BIO307 Lecture 5 (Enzyme Kinetics I)Document11 pagesBIO307 Lecture 5 (Enzyme Kinetics I)Phenyo Mmereki100% (1)
- 1.carbohydrates and Lipid Metabolism-Converted - WatermarkDocument97 pages1.carbohydrates and Lipid Metabolism-Converted - WatermarkJuliyamol JoseNo ratings yet
- Neuron Structure and Function: Active Lecture Question Slides Prepared by Dr. Alan F. Smith, Mercer UniversityDocument105 pagesNeuron Structure and Function: Active Lecture Question Slides Prepared by Dr. Alan F. Smith, Mercer UniversityDenden Gamboa100% (1)
- Michaelis Menten EquationDocument9 pagesMichaelis Menten Equationsadaf zaidiNo ratings yet
- Enzyme Catalysis-Chapter 7 (Part 1)Document22 pagesEnzyme Catalysis-Chapter 7 (Part 1)OmSilence2651No ratings yet
- CSIM2.24 - Signal TransductionDocument6 pagesCSIM2.24 - Signal TransductionAinahMahaniNo ratings yet
- BIO307 Lecture 3 (Introduction To Enzymes)Document13 pagesBIO307 Lecture 3 (Introduction To Enzymes)Phenyo MmerekiNo ratings yet
- Protein Biocatalyst (Enzyme)Document3 pagesProtein Biocatalyst (Enzyme)Khairul RidwanNo ratings yet
- Lecture 5a CatalysisDocument22 pagesLecture 5a CatalysisSandeep ChawlaNo ratings yet
- Protein PurificationDocument7 pagesProtein PurificationArchana BorahNo ratings yet
- Chemical Energetics Notes EditedDocument12 pagesChemical Energetics Notes EditedDaniel Png100% (1)
- Cleavable LinkersDocument12 pagesCleavable LinkersSrinivasa Reddy Telukutla100% (1)
- 1.1 Proteins - Motifs Structural and Functional Domains Protein FamiliesDocument32 pages1.1 Proteins - Motifs Structural and Functional Domains Protein FamilieskinjalkaNo ratings yet
- Dominica State College Biochemistry EnzymesDocument11 pagesDominica State College Biochemistry EnzymesShirlyn GeorgeNo ratings yet
- Applications of Spectroscopic TechniquesDocument20 pagesApplications of Spectroscopic Techniquesamanbioq1No ratings yet
- DNA Structure - ReplicationDocument22 pagesDNA Structure - Replicationapi-3763146100% (1)
- Nucleotide MetabolismDocument89 pagesNucleotide Metabolismfeby ary annasNo ratings yet
- Concise Biochemistry: Fundamental Principles: March 2016Document52 pagesConcise Biochemistry: Fundamental Principles: March 2016Sagar DeshmaniNo ratings yet
- STRAIN IMPROVEMENT TechniquesDocument28 pagesSTRAIN IMPROVEMENT TechniqueselaiyarajaNo ratings yet
- Structure and Function of Bio-Molecules: 9 2. Proteins 13Document62 pagesStructure and Function of Bio-Molecules: 9 2. Proteins 13Alex-Mihai CiubaraNo ratings yet
- Methods of Protein AnalysisDocument41 pagesMethods of Protein AnalysisObin MrazNo ratings yet
- The Kinetics of Enzyme - Catalyzed ReactionsDocument38 pagesThe Kinetics of Enzyme - Catalyzed ReactionsRojan Pradhan100% (1)
- Biochemistry of Kidneys and UrineDocument18 pagesBiochemistry of Kidneys and UrineAndrias PutriNo ratings yet
- Chem 40 Enzyme KineticsDocument85 pagesChem 40 Enzyme KineticsJustine Grace Mariano100% (1)
- Antineoplastic AgentsDocument10 pagesAntineoplastic AgentsMuhamed ArsalanNo ratings yet
- Buffers Notes1Document3 pagesBuffers Notes1Lara MonevaNo ratings yet
- Chapter 2 - Basics of EnzymesDocument40 pagesChapter 2 - Basics of EnzymesSakinah MuhamadNo ratings yet
- Biotin in Metabolism and Its Relationship To Human DiseaseDocument9 pagesBiotin in Metabolism and Its Relationship To Human DiseaseMaikel Ignacio Z. PeñaNo ratings yet
- Topic 5 - Microbial HabitatDocument87 pagesTopic 5 - Microbial HabitatWahyuni IrmalNo ratings yet
- 3-D Structure of ProteinsDocument22 pages3-D Structure of Proteinsraanja2No ratings yet
- Enzymes: A Protein With Catalytic Properties Due To Its Power of Specific ActivationDocument35 pagesEnzymes: A Protein With Catalytic Properties Due To Its Power of Specific ActivationAkash SinghNo ratings yet
- 2.4 Proteins: Essential Idea: Proteins Have A Very Wide Range of Functions in Living OrganismsDocument31 pages2.4 Proteins: Essential Idea: Proteins Have A Very Wide Range of Functions in Living OrganismsMubarak AlNaimiNo ratings yet
- 1.1 Enzymology (Bravo)Document11 pages1.1 Enzymology (Bravo)Arman Carl DulayNo ratings yet
- Lec04 MicroDocument13 pagesLec04 MicroMayurdhvajsinh JadejaNo ratings yet
- 22 CH106 Metabolic Paths For Carbohydrates Timberlake 2ndDocument70 pages22 CH106 Metabolic Paths For Carbohydrates Timberlake 2ndEnrique LiKeNo ratings yet
- Boger CourseDocument477 pagesBoger CourseharrypoutreurNo ratings yet
- 2 Bioenergetics and Oxidative Metabolism IDocument3 pages2 Bioenergetics and Oxidative Metabolism ILinus LiuNo ratings yet
- Chapter 23Document53 pagesChapter 23haririNo ratings yet
- Acids and Bases: Key Concepts of Brønsted-Lowry and Lewis TheoriesDocument27 pagesAcids and Bases: Key Concepts of Brønsted-Lowry and Lewis TheoriesIstiNo ratings yet
- 2.1 Molecules To Metabolism-STUDENTDocument54 pages2.1 Molecules To Metabolism-STUDENTAngel Alexandra SiregarNo ratings yet
- Amino Acids QuestionsDocument5 pagesAmino Acids QuestionsKrishna KumarNo ratings yet
- Chapter 8 - Introduction To MetabolismDocument37 pagesChapter 8 - Introduction To Metabolismprehealthhelp88% (8)
- Transport Processes and Carrier DesignDocument24 pagesTransport Processes and Carrier Designcgckamilshaikh50% (2)
- Enzymes: Protein Catalysts That Increase The Rate of Reactions Without Being Changed in The Overall ProcessDocument49 pagesEnzymes: Protein Catalysts That Increase The Rate of Reactions Without Being Changed in The Overall ProcessGhafoor AzamNo ratings yet
- Electrochemistry Lect Notes CambridgeDocument4 pagesElectrochemistry Lect Notes Cambridgeavatar_75No ratings yet
- Book Solution FoyesDocument15 pagesBook Solution Foyesnorma ireneNo ratings yet
- QUESTION 1 (52 Marks) : Biochemistry 3 BCH 314Document4 pagesQUESTION 1 (52 Marks) : Biochemistry 3 BCH 314victorNo ratings yet
- Second Messenger Signaling SystemsDocument42 pagesSecond Messenger Signaling SystemsMirza Shaharyar BaigNo ratings yet
- Sutherland 1991Document7 pagesSutherland 1991Isal AbdussalamNo ratings yet
- Respiratory Chain & Oxidative Phosphorylation TutorialDocument57 pagesRespiratory Chain & Oxidative Phosphorylation TutorialHanifa AffianiNo ratings yet
- Enzyme Chemistry of Phenolic Compounds: Proceedings of the Plant Phenolics Group Symposium, Liverpool, April 1962From EverandEnzyme Chemistry of Phenolic Compounds: Proceedings of the Plant Phenolics Group Symposium, Liverpool, April 1962J. B. PridhamNo ratings yet
- InterferonsFrom EverandInterferonsThomas MeriganNo ratings yet
- Industrial Relations and Labor LawsDocument6 pagesIndustrial Relations and Labor LawsmengelhuNo ratings yet
- Second Monthly TestDocument4 pagesSecond Monthly TestmengelhuNo ratings yet
- My English Reader Std-III - 2015-1Document32 pagesMy English Reader Std-III - 2015-1mengelhu75% (4)
- Mca EntranceDocument9 pagesMca EntrancemengelhuNo ratings yet
- Office of The Principal Notice No: 600 Date: 27/01/2016: ND TH TH STDocument5 pagesOffice of The Principal Notice No: 600 Date: 27/01/2016: ND TH TH STmengelhuNo ratings yet
- New Microsoft Office Word DocumentDocument2 pagesNew Microsoft Office Word DocumentmengelhuNo ratings yet
- Being A StudentDocument2 pagesBeing A StudentmengelhuNo ratings yet
- Performance ManagementDocument7 pagesPerformance ManagementmengelhuNo ratings yet
- Management Development Approaches (MBHS3Document11 pagesManagement Development Approaches (MBHS3mengelhuNo ratings yet
- Blis 02Document1 pageBlis 02mengelhuNo ratings yet
- Sons of The Soil Theory in RecruitmentDocument1 pageSons of The Soil Theory in Recruitmentmengelhu100% (1)
- No - Date - : To The Controller of Examinations F.M. University, Vyasa Vihar BalasoreDocument2 pagesNo - Date - : To The Controller of Examinations F.M. University, Vyasa Vihar BalasoremengelhuNo ratings yet
- E Assessment: Nit Mca Common Entrance Test (Nimcet) SyllabusDocument3 pagesE Assessment: Nit Mca Common Entrance Test (Nimcet) SyllabusmengelhuNo ratings yet
- PGDCL ProjectDocument5 pagesPGDCL ProjectmengelhuNo ratings yet
- Rural Devlopment Plan:-: Rural Development Is The Process of Improving TheDocument1 pageRural Devlopment Plan:-: Rural Development Is The Process of Improving ThemengelhuNo ratings yet
- SaumyaDocument10 pagesSaumyamengelhuNo ratings yet
- MBA Finance ProjectDocument1 pageMBA Finance ProjectmengelhuNo ratings yet
- Municipal Solid WasteDocument3 pagesMunicipal Solid WastemengelhuNo ratings yet
- VUMC Project Implementation Process GuidelinesDocument48 pagesVUMC Project Implementation Process GuidelinesmengelhuNo ratings yet
- Summing Digits of A NumberDocument1 pageSumming Digits of A NumbermengelhuNo ratings yet
- What Is Human Resource Developmet?: Definitions of HRDDocument4 pagesWhat Is Human Resource Developmet?: Definitions of HRDmengelhuNo ratings yet
- Akash PradhanDocument1 pageAkash PradhanmengelhuNo ratings yet
- Write Note OnDocument2 pagesWrite Note OnmengelhuNo ratings yet
- Masama Aaru Maasama Engeyum Eppothum HD 1080PDocument1 pageMasama Aaru Maasama Engeyum Eppothum HD 1080PmengelhuNo ratings yet
- LetterDocument1 pageLettermengelhuNo ratings yet
- The Request Is Submitted SuccessfullyDocument1 pageThe Request Is Submitted SuccessfullymengelhuNo ratings yet
- 2nd Int (BS) - PKPDocument1 page2nd Int (BS) - PKPmengelhuNo ratings yet
- ApplicationDocument1 pageApplicationmengelhuNo ratings yet
- NAFPDocument1 pageNAFPmengelhuNo ratings yet
- Check UAN StatusDocument1 pageCheck UAN StatusmengelhuNo ratings yet
- Chap. 6B Enzymes: - Introduction To Enzymes - How Enzymes Work - Enzyme Kinetics As An Approach ToDocument28 pagesChap. 6B Enzymes: - Introduction To Enzymes - How Enzymes Work - Enzyme Kinetics As An Approach ToRock SmithNo ratings yet
- Biochemistry ACS Study GuideDocument14 pagesBiochemistry ACS Study GuideLexi TinsleyNo ratings yet
- Nzymes: By: Mrs. Kalaivani Sathish. M. Pharm, Assistant Professor, Pims - PanipatDocument63 pagesNzymes: By: Mrs. Kalaivani Sathish. M. Pharm, Assistant Professor, Pims - Panipaturmila pandeyNo ratings yet
- The Organic Chemistry of Enzyme-Catalysed ReactionsDocument63 pagesThe Organic Chemistry of Enzyme-Catalysed ReactionskunwarskNo ratings yet
- 02 - Enzyme Catalysis PDFDocument42 pages02 - Enzyme Catalysis PDFJavier TorresNo ratings yet
- Biochem Enzyme NotesDocument8 pagesBiochem Enzyme NotesBeatriz NideaNo ratings yet
- EnzymesDocument80 pagesEnzymesThisha MohanNo ratings yet
- Bich411 Enzyme Catalysis PDFDocument7 pagesBich411 Enzyme Catalysis PDFAn TranNo ratings yet
- Effect of Temperature PH Enzyme To Substrate Ratio Substrate Concentration and Time On The Antioxidative Activity of Hydrolysates From Goat Milk Casein by AlcalaseDocument10 pagesEffect of Temperature PH Enzyme To Substrate Ratio Substrate Concentration and Time On The Antioxidative Activity of Hydrolysates From Goat Milk Casein by AlcalaseKhoirul MarifahNo ratings yet
- Biochemistry Past Paper QuestionDocument20 pagesBiochemistry Past Paper QuestionAchraf RabadiNo ratings yet
- Bio Chemistry: Core Unit #1 - Review and IntroductionDocument28 pagesBio Chemistry: Core Unit #1 - Review and Introduction0921pyNo ratings yet
- General Biology 1: Quarter 1 - Week 7-Module 7Document9 pagesGeneral Biology 1: Quarter 1 - Week 7-Module 7Carl Gabriel GravilezNo ratings yet
- Vinylic Carbon, Ambient Nucleophilic and Substrate ReactionDocument11 pagesVinylic Carbon, Ambient Nucleophilic and Substrate Reactiongmanooj_484213209No ratings yet
- Biochemistry Midterm TopicsDocument18 pagesBiochemistry Midterm TopicsAndrea Jersey BuluranNo ratings yet
- BMS131 Lec05Document19 pagesBMS131 Lec05mariam tarekNo ratings yet
- Enzyme Kinetics: Enzyme Kinetics Is The Study of The ChemicalDocument15 pagesEnzyme Kinetics: Enzyme Kinetics Is The Study of The ChemicalDr. SHIVA AITHALNo ratings yet
- C9e Answers Active Reading 08Document7 pagesC9e Answers Active Reading 08melissa_chow_1100% (6)
- Biochem Sample Test-2-2016-AnswersDocument6 pagesBiochem Sample Test-2-2016-Answerssuperfr3shmNo ratings yet
- Biochemistry A Short Course 2Nd Edition Tymoczko Test Bank Full Chapter PDFDocument23 pagesBiochemistry A Short Course 2Nd Edition Tymoczko Test Bank Full Chapter PDFciaramilcahbrpe100% (13)
- Long Answer Questions A LEVEL BIOLOGYDocument205 pagesLong Answer Questions A LEVEL BIOLOGYakil75% (4)
- Sachal Sarmast Coaching AcademyDocument6 pagesSachal Sarmast Coaching AcademySagar Mal MalNo ratings yet
- Theories of Mechanism of EnzymeDocument10 pagesTheories of Mechanism of EnzymeNandini AgrawalNo ratings yet
- Biochem Computational FullDocument390 pagesBiochem Computational Fullrafael_nicolay9196No ratings yet
- 2.5 Enzyme Skeleton NotesDocument8 pages2.5 Enzyme Skeleton Notesadri baigorriNo ratings yet
- Chapter 3: Amino Acids & Peptides: Dr. Bacolod, Chuckie Miguel A. Silliman University Medical SchoolDocument30 pagesChapter 3: Amino Acids & Peptides: Dr. Bacolod, Chuckie Miguel A. Silliman University Medical SchoolThea Cheruve TomentosNo ratings yet
- Enzymes Are Always: Compressed Notes Chapter 4: Biocatalysis Sb025Document7 pagesEnzymes Are Always: Compressed Notes Chapter 4: Biocatalysis Sb025LIM ZHI SHUENNo ratings yet
- Biology Paper 3 Experiment - Activity Enzyme Affected by TemperatureDocument18 pagesBiology Paper 3 Experiment - Activity Enzyme Affected by TemperaturePau Siew LingNo ratings yet
- Chapter 8 Study Guide Ap BioDocument6 pagesChapter 8 Study Guide Ap BioIllan Chichportich100% (1)
- Materi Kuliah Biokimia Stmi Ke 11 (Enzyme Kinetics)Document72 pagesMateri Kuliah Biokimia Stmi Ke 11 (Enzyme Kinetics)soni_abdullahNo ratings yet
- bk978 0 7503 1302 5ch1Document31 pagesbk978 0 7503 1302 5ch1Vaibhav RAJ KUMAR GUPTANo ratings yet