Professional Documents
Culture Documents
Lectura Examen Final
Uploaded by
Anonymous M5KvpjCuCopyright
Available Formats
Share this document
Did you find this document useful?
Is this content inappropriate?
Report this DocumentCopyright:
Available Formats
Lectura Examen Final
Uploaded by
Anonymous M5KvpjCuCopyright:
Available Formats
Captulo
La clula: evolucin y desarrollo
de los sistemas celulares
Jorge Joel Reyes Mndez
El objetivo de la biologa celular y molecular moderna es interpretar las propiedades
de los organismos mediante la estructura de sus molculas constituyentes.
FRANCOIS JACOB, 1973, en The logic of life.
Contenido
Objetivos
Introduccin
Desde las molculas hasta la primera
clula
Formacin de molculas sencillas en las
condiciones prebiticas
Sntesis de los polinucletidos
Seleccin natural de las molculas
automultiplicadoras
Molculas de RNA como catalizadores
Transferencia de informacin desde
los polinucletidos a los
polipptidos
Formacin de la membrana
Primeras clulas
De la clula procariota a la eucariota
Clulas procariotas
Reacciones metablicas
Fotosntesis
Respiracin
Gluclisis
La clula eucariota
Teora actual de la endosimbiosis como
origen de la clula eucariota
Estructura celular
Transporte a travs de membranas
Transporte pasivo
Difusin simple
Transporte activo
Citosol y citoesqueleto
Microtbulos
Microfilamentos
Filamentos intermedios
Distribucin de los filamentos del
citoesqueleto en el citoplasma
Retculo endoplsmico
Aparato o complejo de Golgi
Lisosomas
Mitocondria
Cloroplasto
Fotosistemas
Ncleo
Cromatina y cromosomas
El gen desnudo
Reproduccin de los virus
Viroides
Priones
Captulo 1
La clula: evolucin y desarrollo de los sistemas celulares
Objetivos
Al nalizar el captulo, el lector podr identicar las teoras
actuales sobre el origen de los sistemas precelulares y de la
vida; interpretar y conceptualizar los procesos evolutivos que
llevaron a la aparicin de los procariotas; sealar los fundamentos de la teora endosimbitica del origen de las clulas
eucariotas; identicar las diferencias fundamentales entre una
clula procariota y una clula eucariota y describir el proceso
de sntesis de protenas en sus partes fundamentales.
Introduccin
Hace aproximadamente 4 700 millones de aos, segn los
clculos ms recientes, en uno de los brazos de la galaxia,
una nube de gas y polvo se contrajo para formar un disco.
Durante dicha contraccin, esta nube aument su velocidad
angular. Hubo un momento en el que el centro del disco fue
tan denso, masivo y caliente que fue posible que se llevaran
a cabo reacciones nucleares en su interior. Este hecho lo
convirti en una estrella, el Sol. Posteriormente, parte de las
partculas de polvo y gas restantes se unieron para dar origen
a los planetas.
Setecientos millones de aos despus, es decir, hace
cuatro mil millones de aos, la Tierra completaba un giro
alrededor de su eje en 18 horas. El joven Sol brillaba con slo
el 70% de su energa actual. La atmsfera no contena oxgeno
libre. Haba pocos sonidos, salvo el ruido del viento, el de las
erupciones volcnicas, el silbido de la lava al hacer contacto
con el agua y las explosiones de los meteoritos al chocar contra
la corteza terrestre. No haba plantas, animales, bacterias, ni
virus. A partir de este punto inicial, la opinin de los cientcos vara ampliamente: no se sabe con exactitud cmo era
el planeta Tierra cuando inici la vida. Las rocas que podran
brindar esta informacin se deformaron hace mucho tiempo
a causa de los movimientos de la corteza terrestre.
Stanley L. Miller arma que los compuestos orgnicos
se organizaron en algo que poda automultiplicarse. Son
varios los escenarios posibles. A continuacin se exponen
los ms recientes puntos de vista de cientcos destacados,
estudiosos del tema.
Aunque los acontecimientos que originaron la vida en
la Tierra an son misteriosos, los cientcos exploran desde
hace algunos aos pistas nuevas que se revelan prometedoras.
El qumico norteamericano Stanley Miller haba efectuado
en los aos cincuenta del siglo XX los primeros experimentos
de reproduccin de una sopa primitiva en la que se habran
sintetizado las molculas que constituyen la materia viva. En
la actualidad, esta hiptesis se considera improbable, y parece
ms verosmil que dichas molculas se hayan formado en la
supercie de determinados minerales. Segn este punto de
vista, uno de los problemas fundamentales es saber cmo
surgieron las molculas capaces de automultiplicarse en ausencia de enzimas, condicin indispensable para que se pueda
transmitir la informacin gentica.
Se sabe que todos los seres vivos estn constituidos por clulas (pequeos compartimientos rodeados con una membrana
y rellenos de una solucin de sustancias qumicas). Las formas
ms simples de vida son clulas solitarias que se reproducen
dividindose en dos. Los organismos restantes, incluso el ser
humano, constan de agrupaciones de clulas que efectan
determinadas tareas y que se comunican con otros grupos de
clulas mediante procedimientos de seales complejos.
Todos los organismos y todas las clulas que los constituyen proceden de un antecesor comn, pero se desconocen
los procesos intermedios. Uno de estos procesos, denominado evolucin, tiene lugar por medio de dos mecanismos
bien denidos:
la variacin mediante la cual se produce en forma aleatoria
un cambio en la informacin gentica
la seleccin en favor de los individuos que estn mejor
preparados para sobrevivir y reproducirse
Por otro lado, se sabe que ms que una curiosidad biolgica, la endosimbiosis es uno de los motores ms poderosos
de la evolucin del mundo vivo. La endosimbiosis crea con
rapidez organismos quimricos que engendran nuevos linajes,
acerca a los socios y favorece transferencias masivas de genes
que crean genomas tambin quimricos: el genoma nuclear
contiene, por consiguiente, genes eucariotas, pero tambin
genes de origen bacteriano procedente de las mitocondrias
o de los plastos, con los que coexiste. Estos acontecimientos
explican tal vez los saltos evolutivos fundamentales que dieron
origen a los grandes linajes del mundo viviente y moldearon
la diversidad biolgica actual. El mismo hombre puede considerarse como una comunidad simbitica, que est formada
por el citoplasma eucariota y las mitocondrias.
El hombre es, en realidad, un conglomerado de bacterias,
espiroquetas y virus diversos. El producto acabado de una
industriosa colonia de bacterias simbiticas que lo armaron,
de la misma forma que el hombre construye su hogar y sus
ciudades.
Este captulo trata de la progresin desde las molculas
hasta los organismos multicelulares. En seguida se estudian
las estructuras fundamentales de las clulas eucariticas (clulas que disponen de una anatoma intracelular estructurada
a diferencia de las clulas procariticas que carecen de estas
estructuras).
Captulo 1 La clula: evolucin y desarrollo de los sistemas celulares
Desde las mleculas hasta
la primera clula
Formacin de molculas sencillas
en las condiciones prebiticas
En 1970 se descubri la existencia de cido frmico en la
regin norte de la galaxia Sagittarius B2, y la conrmaron
radioastrnomos de la Universidad de Illinois en 1999. Por
otra parte, en 1996, cientcos de esta misma universidad
encontraron cido actico en esta misma nube de gas. El cido
actico es una molcula slo un paso ms corta que la glicina,
el aminocido ms sencillo, cuya existencia en el espacio no se
ha conrmado todava. De hecho, para la formacin de glicina
tan slo habra que combinar cido actico con amoniaco, cuya
presencia en el espacio se conoce, a su vez, desde 1969.
Se descubrieron alrededor de 120 molculas diferentes en
esas nubes. Algunas de ellas tienen ocho o ms tomos, como
el glicoaldehdo, aunque la mayora contiene menos.
En el ao 2000, un equipo de cientcos formado por
Jan Hollis (NASA/Goddard), Frank J. Lovas (Universidad de
Illinois) y Philip R. Jewell (NRAO/Green Bank) descubri
glicoaldehdo en una nube gigante de gas y polvo situada a
una distancia de alrededor de 26 000 aos luz de la Tierra,
a unos 300 aos luz del centro de la galaxia, la Va Lctea,
por medio del radiotelescopio de 12 metros de la Fundacin
Nacional para la Ciencia, en Kitt Peak, a 80 Km de Tucson,
Arizona. Tales nubes, a menudo con un tamao de varios aos
luz, estn compuestas por material a partir del cual se forman
nuevas estrellas. Aunque su densidad es muy pequea, se trata
de zonas en las que se suceden complejas reacciones qumicas
a lo largo de cientos de miles o de millones de aos.
El glicoaldehdo (o glicolaldehdo, fig. 1-1) es una
molcula de ocho tomos cuya frmula es C2H4O2 (HOCH2CHO). Se trata de un aldehdo con dos tomos de carbono, el
cual tambin se puede incluir en el grupo de las aldosas ms
sencillas, las biosas, carbohidratos (azcares) con un tomo
de carbono menos que las triosas. Se puede combinar con
otras molculas para formar azcares ms complejos, como
ribosa o glucosa.
OH
O
C
H
C
H
OH
O
C
H
H
CH2OH
Figura 1-1. Izquierda: glicoaldehdo; derecha: gliceraldehdo, una triosa.
El descubrimiento de esta molcula de azcar en una nube
a partir de la cual se forman las estrellas incrementa la probabilidad de que los precursores qumicos de la vida se originen
en estas nubes, mucho antes de que los planetas se creen alrededor de las estrellas. Dicho descubrimiento puede ser una
clave importante para saber cmo surgi la vida en la Tierra.
Las condiciones existentes en las nubes interestelares pueden en
ciertos casos ser similares a las condiciones que privaban en la
Tierra en sus primeras etapas, por lo que su estudio ayudara a
los cientcos a comprender cmo se formaron las biomolculas en las primeras etapas de la historia del planeta Tierra (g.
1-2). Por otro lado, algunos cientcos sugieren que el paso de
cometas podra haber sembrado molculas complejas, hechas
de material proveniente de nubes interestelares, en la Tierra.
Aunque no se conoce con exactitud la composicin de
la atmsfera primitiva, se cree que adems de vapor de agua
contena dixido de carbono, metano o amoniaco e hidrgeno.
Tambin se supone que haba una intensa actividad volcnica
con temperaturas elevadas, lluvias torrenciales y continuas
tormentas elctricas. Adems, no haba oxgeno libre, ni
capa de ozono que protegiera a la Tierra contra la radiacin
ultravioleta. En estas condiciones ambientales se formaron
seguramente las primeras molculas orgnicas, cianuro de
hidrgeno, formaldehdo, cido actico, etc., que habran
reaccionado con facilidad para formar los cuatro componentes fundamentales de la vida:
aminocidos
azcares o glcidos
lpidos o grasas
nucletidos
Esta hiptesis se comprob mediante experimentos de
laboratorio. En 1953 Stanley L. Miller mostr que si una
mezcla de gases, como metano, amoniaco, dixido de carbono
e hidrgeno se calentaba con vapor de agua y se someta a
descargas elctricas y a radiacin ultravioleta durante varias
semanas, se formaban los cuatro tipos de molculas mencionados (g. 1-3).
Sntesis de los polinucletidos
Las molculas sencillas, como los aminocidos o los nucletidos, se pueden agrupar para formar grandes polmeros. Un
aminocido puede unirse a otro mediante un enlace peptdico
y un nucletido puede unirse con otro por medio de un enlace fosfodister. La repeticin de estas reacciones conduce,
respectivamente, a un polipptido y a un polinucletido,
componentes fundamentales de las protenas y de los cidos
ribonucleico y desoxirribonucleico. Las protenas se forman
a partir de un conjunto de 20 aminocidos, mientras que los
cidos nucleicos estn compuestos exclusivamente por cuatro
tipos de nucletidos (g. 1-4).
Captulo 1
Hace
1 500
millones
de aos
La clula: evolucin y desarrollo de los sistemas celulares
Atmsfera
Big Bang
4 500
4 000
Formacin
de galaxia
Geosfera
Escape
del H2
500
Enfriamiento
de corteza
y atmsfera
Gases
volcnicos
Vida
en tierra
firme
Evolucin
qumica
Ocanos
Acumulacin
de materia
orgnica
Aparicin
de cmulos
globulares
10 000
Biosfera
Cdigo
gentico
Gluclisis
400
Evolucin
prebitica
Protoclulas
3 000
Desaparicin
de las molculas
orgnicas
del ambiente
300
Algas
2 000
Fotosntesis
Formacin del
sistema solar
4 500
Evolucin
biolgica
Acumulacin
de oxgeno
200
Respiracin
Multicelularidad
Formacin de
la Tierra
Aparicin
de los
dinosaurios
Aparicin
de los
mamiferos
1 000
100
Capa
de ozono
Extincin de
dinosaurios
500
Animales
y vegetales
modernos
Oxgeno
actual
0
Hombre
primitivo
Figura 1-2. Escala del tiempo geolgico y algunos de los principales hechos del origen de la vida.
+
CH4
NH3
CO2
H2
Descarga
elctrica
Cromatografa
sobre papel
Agua
Trampa
Calor
Figura 1-3. Experimento de Miller.
Formaldehdo
cido frmico
Cianuro de hidrgeno
cido actico
cido lctico
Glicina
Alanina
Urea
cido asprtico
Captulo 1 La clula: evolucin y desarrollo de los sistemas celulares
G
C
G
C
H2O
Figura 1-4. Los cuatro tipos de nucletidos pueden experimentar una polimerizacin espontnea con
prdida de agua. El resultado es una mezcla de polinucletidos de longitud y orden aleatorios.
Los polmeros primitivos pueden haberse formado de
varias maneras, por ejemplo, al calentarse los constituyentes
orgnicos en seco o al catalizar la reaccin altas concentraciones de polifosfatos inorgnicos. En condiciones de laboratorio, el resultado son polmeros de longitud variable, cuyos
componentes se acomodan en forma aleatoria (g. 1-5). Pero
una vez que se ha formado un polmero, ste puede inuir
en la formacin de uno nuevo, ya que puede actuar como
molde en una nueva reaccin de polimerizacin. En efecto,
los nucletidos tienen la propiedad de emparejarse mediante
la formacin de enlaces de hidrgeno, de tal manera que un
polmero (p. ej., el poli U) induce la formacin del polmero
complementario (poli A) (g. 1-6).
Probablemente el emparejamiento especfico de los
nuclotidos (bases) desempe un papel decisivo en el surgimiento de la vida. Considrese, por ejemplo, un fragmento
de RNA que contiene las cuatro bases: adenina, A, uracilo, U,
citosina, C, y guanina, G. Gracias a la complementariedad de
las bases, si este RNA se aade a una mezcla de nucletidos
activados en condiciones que favorezcan la polimerizacin,
se crear un RNA complementario del primero. Es decir,
las muevas molculas son como el vaciado del molde
G
C
G
H2O
U
C
A
A
G
A
original en donde A ha sido sustituido por U, C por G y
as sucesivamente. La informacin contenida en la secuencia
original se preserva, de algn modo, en las copias, ya que en
una segunda vuelta de polimerizacin, stas generan RNA
idntico al original. De esta forma, la informacin gentica
contenida en la secuencia de nucletidos pasa de generacin
en generacin mediante este proceso de emparejamiento de
bases complementarias.
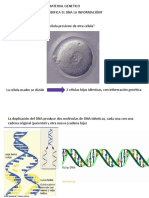
Los mecanismos de duplicacin requieren la presencia
de un catalizador. Sin ste, la reaccin es lenta y poco eciente, y otras reacciones secundarias podran interferir, lo cual
impedira la formacin de rplicas exactas. En la actualidad,
una protena cataliza esta reaccin, la polimerasa de DNA
inexistente en la sopa prebitica. En los tiempos primitivos,
iones metlicos y minerales como la arcilla podran haber
cumplido la funcin de catlisis. Pero, lo que es ms importante, el propio RNA pudo haber actuado como catalizador.
En efecto, el RNA, adems de funcionar como molde para
la duplicacin, tiene la posibilidad de formar plegamientos
intramoleculares que originan supercies complejas, las cuales
catalizan reacciones especcas. Posteriormente se tratan los
mecanismos propuestos a este respecto.
G
C
U
C
Figura 1-5. Los nucletidos muestran preferencias al formar parejas: A con U, C con G, mediante la
formacin de puentes de hidrgeno. De esta forma, un polinucletido sirve de molde para un polinu-
Captulo 1
La clula: evolucin y desarrollo de los sistemas celulares
Paso1
Paso 2
LA SECUENCIA ORIGINAL
FORMA
LA SECUENCIA COMPLEMENTARIA
Figura 1-6.
LA SECUENCIA COMPLEMENTARIA
FORMA
LA SECUENCIA ORIGINAL
Es inevitable que, en los procesos de copia como el ya descrito,
se produzcan errores y que una base sea sustituida por otra en
alguna copia. De este modo, a partir de una molcula original se pueden generar multitud de variaciones. Ahora bien,
estas molculas tienen propiedades qumicas especcas. Una
secuencia determinada de bases conere un comportamiento
diferente a la molcula. En particular, la secuencia de bases
establece cmo puede plegarse la molcula sobre s misma
cuando est en solucin. En efecto, de la misma manera que
en la duplicacin, las bases de una secuencia forman parejas
con las bases complementarias libres que estn en solucin;
en una molcula nica de RNA de gran tamao se generan
parejas de secuencias complementarias que existen dentro del
polmero. Entonces, por ejemplo, si dentro de una molcula
de RNA existe la secuencia AAAAA, sta podr emparejarse con una secuencia UUUUU que exista en otra parte
de la molcula (g. 1-7). Este tipo de vnculo produce un
plegamiento caracterstico que depende slo de la secuencia
de nucletidos.
La estructura tridimensional de un polinucletido estipula su estabilidad qumica, sus acciones sobre otras molculas
y su capacidad para copiarse, de tal forma que no todas las
molculas son igualmente viables en una mezcla. Mediante
estudios de laboratorio se demostr que algunas molculas
de un sistema replicante de RNA experimentan una seleccin natural: predominan algunas secuencias favorables, que
dependen de las condiciones experimentales.
Por lo tanto, la molcula de RNA tiene dos propiedades
fundamentales:
Seleccin natural de las
molculas automultiplicadoras
lleva informacin codicada en una secuencia de nucletidos, orden que puede transferir mediante copia
tiene una estructura tridimensional nica que determina
cmo interaccionar con otras molculas y cmo responder
a las condiciones del ambiente
Estas dos propiedades informativa y funcional son
fundamentales en la evolucin. La informacin codicada del
RNA es igual al genotipo informacin hereditaria, en
tanto que la forma expresada es igual al fenotipo expresin
de la informacin gentica.
Molculas de RNA como catalizadores
La seleccin natural depende del ambiente. Para una
molcula de RNA replicante es importante la presencia de
otras molculas de RNA en el medio. Adems de su papel
como moldes, algunas molculas de RNA con plegamientos
determinados pueden actuar como catalizadores de algunas
reacciones qumicas, incluso la rotura y la formacin de en-
G AA
A
CU
GAU
A
G
A
G
G
AU G
CUU
U C
C U
U G
C G
C A
C
C
G
A
C
C
U
G
G
UA C A
Figura 1-7. El emparejamiento intramolecular de nucletidos produce un plegamiento de la molcula de RNA.
Captulo 1 La clula: evolucin y desarrollo de los sistemas celulares
laces covalentes y enlaces entre nucletidos. Por ejemplo, un
RNA especializado puede catalizar el corte de otro RNA o
de s mismo en una secuencia determinada. Cada una de las
reacciones qumicas que cataliza el RNA cataltico depende
de una conformacin o agrupamiento especco de los tomos
que constituyen su supercie, lo cual hace que uno o varios
nucletidos sean altamente reactivos.
Estas reacciones catalticas pudieron tener una importancia fundamental en la sopa prebitica. En efecto, supngase
que una molcula de RNA es capaz de catalizar el proceso
de polimerizacin a partir de una molcula cualquiera de
RNA como molde. Esta molcula cataltica har copias de s
misma, pero tambin de cualquier otro RNA que se encuentre en la proximidad, las cuales pueden ayudar, a su vez, a la
reproduccin o supervivencia de la primera. De esta manera,
la especializacin de diferentes RNA constituira un sistema
eciente capaz de copiarse y de sobrevivir (g. 1-8).
Transferencia de informacin desde
los polinucletidos a los polipptidos
Se supone que hace entre 3.5 y 4 mil millones de aos comenz la evolucin de las molculas automultiplicadoras o
autorreplicantes de RNA. De la misma manera que ahora los
organismos compiten para sobrevivir, sistemas compuestos de
varios tipos de RNA, como los de la gura 1-9, competan
por los precursores que les servan para automultiplicarse. Su
capacidad para sobrevivir dependa de la exactitud y eciencia
de la copia, as como de la estabilidad de la misma.
Sin embargo, aunque las molculas de RNA tienen la
aptitud de codicar y mantener informacin con slo cuatro
nucletidos o bases, no son muy capaces de catalizar reacciones
complejas. Los polipptidos, que estn compuestos por ms
aminocidos, son mucho ms capaces de crear puntos activos
catalticos por medio del plegamiento de sus cadenas. Incluso
algunos polipptidos creados al azar uniendo ciertos aminocidos pueden mostrar capacidades catalticas superiores a las
del mismo RNA para la duplicacin de ste.
Por consiguiente, cualquier sistema de RNA con un
polipptido cataltico habra tenido mayores probabilidades
de sobrevivir y, de igual manera, un polinucletido capaz de
estimular la sntesis de un polipptido cataltico habra presentado grandes ventajas sobre los dems.
Cmo pudieron los polinucletidos ejecutar tal tarea?
Cmo pudo la informacin codicada en sus secuencias
especicar el orden de diferentes tipos de polmeros? Se sabe
que los polinucletidos pueden actuar como catalizadores para
unir diferentes aminocidos. En la actualidad, la sntesis de
protenas se lleva a cabo mediante un conjunto de sistemas
en los que desempean un papel importante tipos distintos
de RNA. Sin embargo, en el proceso intervienen tambin
diferentes protenas previamente sintetizadas.
replicacin
catlisis
RNA cataltico que une
nucletidos para formar
una nueva molcula con
la misma secuencia y
forma
una copia de s mismo
RNA cataltico que une
nucletidos para formar:
una copia de otro RNA
una copia de otro RNA
Figura 1-8.
En resumen, la sntesis de protenas tiene lugar de la
forma siguiente:
una molcula de RNA contiene la informacin gentica
para sintetizar un determinado polipptido, informacin
que est codicada
otras molculas de RNA actan como portadoras al unirse
a un tipo determinado de aminocido.
Estos dos tipos de molculas de RNA tienen secuencias
complementarias, por lo que las instrucciones que estn
contenidas en la cadena que posee el cdigo se transere al
RNA que aade el aminocido a la cadena polipeptdica en
formacin. Probablemente los precursores de estos dos tipos
de molculas de RNA dirigieron la primera sntesis de protenas sin la ayuda de otras protenas.
En la actualidad, estos procesos de ensamble de aminocidos tienen lugar en la supercie de los ribosomas, partculas
complejas constituidas por varias molculas de RNA de un
nuevo tipo y por lo menos de 50 protenas. Este RNA tiene
un papel cataltico fundamental y constituye el 60% del
Captulo 1
La clula: evolucin y desarrollo de los sistemas celulares
RNA cataltico
una copia de otro RNA
RNA cataltico
una copia de otro RNA
RNA codificado molde
para una protena
RNA transportador
de aminocidos
RNA codificado molde para una protena
RNA transportador
de aminocidos
Figura 1-9.
ribosoma. Esto destaca, al menos desde un punto de vista
evolutivo, la importancia del RNA en la primitiva sntesis
dirigida de protenas.
De esta forma, el RNA fue capaz de ir creando herramientas en forma de protenas para lograr una sntesis
cada vez ms eciente de s mismo y de nuevas molculas de
polinucletidos y de polipptidos.
La sntesis de protenas bajo la direccin de un RNA
requiri la evolucin de un cdigo, el cdigo gentico, por medio
del cual determinadas secuencias de nucletidos especican el
orden en que se debe acomodar los aminocidos que forman
Prote na naciente
la protena. Este cdigo est formado por conjuntos de tres
nucletidos o tripletes, cada uno de los cuales corresponde
a un determinado aminocido. Este cdigo parece haber
sido seleccionado aleatoriamente y est presente en todas las
formas de vida, lo que hace pensar en que todas las clulas
actuales descienden de una misma lnea celular primitiva que
surgi del perfeccionamiento del mecanismo de la sntesis de
protenas.
Una vez que la evolucin de los cidos nucleicos lleg
al punto de disponer de estas molculas proteicas catalticas,
llamadas enzimas, la proliferacin del sistema replicante se
Captulo 1 La clula: evolucin y desarrollo de los sistemas celulares
aceler en gran medida, como ocurre ahora con algunos virus
de algunas bacterias, los cuales se apoderan de la maquinaria
de la clula una vez que entran en ella y dirigen la sntesis de
RNA para automultiplicarse en forma explosiva.
Recientemente dos equipos, uno en Alemania y el otro en
California, han puesto a punto sistemas biomimticos, es decir,
sistemas en los que se multiplican in vitro cidos nucleicos, por
lo cual permiten estudiar este problema. Los cidos nucleicos
DNA y RNA contienen la informacin gentica. Se trata de
polmeros de nucletidos, los cuales se dividen, en funcin
de su estructura qumica, en purinas (adenina y guanina) y
pirimidinas (citosina, timina y uracilo). Los cidos nucleicos
se ligan por medio de enlaces de hidrgeno especcos entre
los nucletidos y forman unas estructuras en doble hlice: estas
uniones especcas son la base fsica de la herencia. Uno de los
escenarios verosmiles para explicar la aparicin de molculas
portadoras de la informacin gentica es el de un mundo
RNA, que propuso el bioqumico norteamericano W. Gilbert
en 1986. Segn esta hiptesis, el RNA fue el primero que se
habra formado, gracias al encadenamiento de nucletidos,
y habra desempeado, en las clulas primitivas, el papel de
DNA y el de protenas, debido en particular a que posee una
actividad cataltica. Luego, en las clulas ms evolucionadas,
el DNA, qumicamente ms estable que el RNA, se habra
encargado de la conservacin de la informacin gentica, y la
actividad cataltica se habra transferido sobre todo a las protenas. El RNA es actualmente el enlace entre los dos, en forma
de RNA mensajero y como constituyente fundamental de los
ribosomas, que son las mquinas que fabrican protenas.
J. Ferris y sus colaboradores, del Instituto Politcnico
Rensselaer, en el Estado de Nueva York, establecieron al
nal de 1993 que se forman pequeos polmeros de nucletidos (oligonucletidos) por medio de catlisis no biolgica. El
dispositivo consiste en una disolucin acuosa de nucletidos,
vueltos articialmente ms reactivos que los nucletidos naturales, agitada en presencia de montmorillonita, con un pH
igual a 8. La montmorillonita es una arcilla natural cida, que
se utiliza de ordinario en qumica orgnica como catalizador, a
la cual se jan los nucletidos antes de reaccionar para formar
pequeos polmeros. Se han detectado oligonucletidos que
constan hasta de once elementos. Es interesante sealar que,
en los cidos nucleicos que se forman, la mayor parte de los
nucletidos se une por medio de enlaces idnticos a los que
se encuentran en los RNA y DNA biolgicos. Por tanto, es
posible la formacin inicial de pequeos polmeros de nucletidos segn un esquema sencillo. No obstante, para validar la
hiptesis del mundo RNA tambin se tiene que encontrar
un mecanismo posible para duplicar los cidos nucleicos sin
enzimas. El qumico norteamericano L. Orgel, pionero de los
estudios sobre la replicacin in vitro de los cidos nucleicos,
demostr, en 1983, que los nucletidos se unen para formar la
hebra complementaria de un cido nucleico rico en pirimidinas.
En cambio, un cido nucleico rico en purinas no puede dirigir la
formacin de la hebra complementaria. Adems, las dos hebras
de una doble hlice de DNA estn enlazadas fuertemente, por
lo que no pueden reiniciar de modo espontneo el proceso y
por eso no hay duplicacin.
Con el n de superar estas dicultades, es necesario utilizar pequeos cidos nucleicos, en vez de simples nucletidos.
Hace poco tiempo, dos qumicos californianos, T. Li y K.
Nicolaou del Instituto de Investigacin Scripps, partieron de
una doble hlice de DNA clsica, del tipo WatsonCrick
(vase g. 1-10A). Aadieron dos oligonucletidos complementarios de una de las hebras de la doble hlice, y los jaron a la doble hlice con lo cual se form una hlice triple.
Luego separaron la hebra recin formada de la triple hlice.
La hebra recin formada dirige, a su vez, el ensamble de los
oligonucletidos que le son complementarios. Se obtiene de
este modo una nueva doble hlice idntica a la doble hlice
inicial (g. 1-10B). Se duplic el DNA. Tambin se examin
la delidad de este proceso de duplicacin. Para ello se utilizaron oligonucletidos en los que uno o varios nucletidos
no corresponden a la doble hlice inicial. Se constata en este
caso que con un solo error de emparejamiento la triple hlice
se forma con ms dicultad, y que no llega a establecerse del
todo cuando hay dos o tres errores. Por tanto, en este sistema
G C
G C
A
T
esqueleto
azcar
fosfato
par de
bases
G C
T A
base
nitrogenada
G C
T A
A
GC
A
Figura 1-10A. Doble hlice de DNA clsica.
10
Captulo 1
La clula: evolucin y desarrollo de los sistemas celulares
REPLICACIN BASADA EN UNA CATLISIS CRUZADA
DE OLIGONUCLETIDOS, DIRIGIDA POR UNA HEBRA MODELO
CICLO DE AMPLIFICACIN POR UNA TRIPLE HLICE
doble hlice inicial (Crick y Watson)
AUTOCOMPLEMENTARIEDAD
C CG C GG
C CGCGG
CCG + C GG
CG G C C G
C GG + C CG
CGGC CG
CCGCGG
C G GC CG
CGG C C G
CCGCC G
(emparejamiento
de Hoogsteen)
triple hlice
agente de condensacin
COMPLEMENTARIEDAD CRUZADA
C GG CGG
CCG +
CCG
CGG +
CGG
C CG C C G
C CG CC G
C G G C GG
disociacin
C C GC CG
C G G C GG
C G G C GG
C C GC CG
copia de la doble
hlice inicial
agente de condensacin
Figura 1-10B.
tambin se pueden copiar algunas secuencias mutadas, pero
con una ecacia menor.
Unos qumicos de la Universidad de Friburgo crearon
paralelamente otro sistema automultiplicador. El equipo de G.
van Kiedrovski estudi la reproduccin competitiva de cuatro
pequeos cidos nucleicos a partir de dos oligonucletidos.
Dos de estos cidos nucleicos dirigen la formacin de productos autocomplementarios; los otros dos, la de productos
complementarios (vase el lado izquierdo del esquema (BA:
vericar a qu gura corresponde)). Al analizar las velocidades
relativas de sntesis de los cuatro cidos nucleicos establecieron
que su copia es un sistema autocataltico, hasta en el caso ms
ordinario de una simple complementariedad de los fragmentos
de cidos nucleicos con la hebra modelo, es decir, independientemente de la secuencia que se duplica.
Resuelven estos dos modelos el problema de la duplicacin del RNA prebitico? El primero propone un
mecanismo sencillo para la multiplicacin del DNA, y el
segundo muestra que la duplicacin del DNA es un proceso
autocataltico cualquiera que sea la secuencia de DNA.
Hay que sealar que en los dos casos, los oligonucletidos
utilizados estn activados, es decir, eran ms reactivos que los
nucletidos naturales, y que se aadieron agentes acopladores
en el momento apropiado: los qumicos ayudan a la automultiplicacin. Por otra parte, el cido nucleico que se estudi
en ambos casos es DNA, que es ms fcil de sintetizar que el
RNA, aunque se supone que este ltimo apareci primero.
Adems, se sintetizaron los oligonucletidos y, luego, se
aadieron al medio reactivo, aunque se habran podido formar de modo natural gracias a la accin cataltica de algunas
rocas como la montmorillonita. Por paradjico que parezca,
no se tiene ninguna idea de la forma en que surgieron los
nucletidos mismos.
Segn J. Ferris, si procesos como los descritos aqu sucedieron para dar origen a la vida en la Tierra, estos nuevos
resultados prueban sobre todo que el problema de la duplicacin
de los cidos nucleicos fuera de una clula tiene solucin, pero
tambin que estaban presentes otros catalizadores distintos del
RNA. Sin embargo, es posible imaginar que fragmentos de
cidos nucleicos, formados por catlisis mineral, se condensaron
a veces en fragmentos ms grandes. De manera gradual habran
adquirido las caractersticas necesarias para hacer vivir el mundo
del RNA. Las primeras clulas vivas habran aparecido al nal
de esta evolucin.
Formacin de la membrana
Es probable que uno de los acontecimientos cruciales en la
evolucin desde los sistemas automultiplicadores hasta la
primera clula haya sido la formacin de una membrana. La
necesidad de mantener una alta concentracin de protenas
Captulo 1 La clula: evolucin y desarrollo de los sistemas celulares
SIN MEMBRANA
11
En una mezcla de RNA
autoreplicantes, un RNA capaz
de producir una protena til
comparte esta protena con otras
especies de RNA competidoras
Enzima
primitiva
punto
activado
Fosfolpidos
ciclo replicante
ACEITE
CON MEMBRANA
AGUA
Enzima
primitiva
membrana
Cuando el RNA y la protena cataltica estn
encerrados en una membrana lipdica, el
sistema aprovecha la enzima en exclusiva.
Figura 1-11.
en las proximidades de un sistema automultiplicador de RNA
y de evitar que otros sistemas vecinos se beneciaran con las
propiedades catalticas de estas protenas (g. 1-11) origin
el paso siguiente de la evolucin: aislar el sistema de RNA
automultiplicador junto con sus protenas catalticas mediante
una membrana para que slo el RNA que las generaba las
pudiera utilizar.
Existen unas molculas, llamadas anpticas, que contienen una parte hidrla (soluble en agua) y una parte lipla (insoluble en agua) que se prestan especialmente para la
formacin de tales membranas. Estas molculas se alinean en
la interfaz aceiteagua para formar una vescula monocapa
(g. 1-12A); cuando se colocan en agua se unen entre s de
manera espontnea y forman unas pequeas vesculas cerradas
bicapa por lo que el contenido acuoso queda aislado del medio
exterior (g. 1-12B).
A las clulas de hoy las rodean membranas que estn
formadas, casi del todo, por fosfolpidos que adoptan dicha
conguracin de bicapas, cuyo ancho mide unos 5 nm y que
se ven a la perfeccin con ayuda del microscopio electrnico.
monocapa de fosfolpidos
Figura 1-12A. Las molculas anfipticas como los fosfolpidos forman de modo espontneo una monocapa en una
interfaz aceite-agua.
Estas membranas aslan las clulas del medio externo hostil
y limitan el espacio ocupado por la clula.
Se ignora en qu momento de la evolucin se form
la primera clula. Puede haberse originado en el momento
en que las primeras molculas de fosfolpidos rodearon un
ACEITE
AGUA
AGUA
AGUA
Figura 1-12B. Los fosfolpidos, cuando se dispersan en
agua, forman espontneamente vesculas monocapa en la
interfaz aceite-agua o vesculas bicapa con las cabezas hidrfilas enfrentando al agua.
12
Captulo 1
La clula: evolucin y desarrollo de los sistemas celulares
sistema automultiplicador de RNA con una vescula bicapa.
Sin embargo, parece ms probable que antes de que estos
sistemas primitivos evolucionaran a clulas tuvo que ponerse
en marcha la sntesis de protenas.
Primeras clulas
Todo lo expuesto antes es puramente especulativo ya que no
existen fsiles de las clulas primitivas. Se supone que la sntesis prebitica de pequeas molculas, la formacin de RNA
autocataltico y la traduccin de las secuencias de RNA a los
primeros polipptidos catalticos tuvieron lugar hace 3 500 o
4 000 millones de aos.
Las primeras clulas debieron ser mucho menos complejas que las clulas ms simples que hoy se conocen, los micoplasmas. stos son pequeos organismos parecidos a bacterias,
parsitos de clulas animales o vegetales. Tienen un tamao
de 0.3 a 0.5 mm y organizan la sntesis de unas 750 protenas
entre catalticas y estructurales, algunas de las cuales estn en
el interior de la clula, otras estn alojadas en la membrana. A
pesar de su simplicidad, los micoplasmas sintetizan todas las
molculas necesarias para su conservacin y reproduccin.
Las clulas primitivas tenan una diferencia importante
con el micoplasma actual: la informacin gentica de este
ltimo est escrita en molculas de DNA y en el caso de la
clula primitiva se utilizaba seguramente RNA. Ambos tipos
de cidos nucleicos estn presentes en la clula de hoy, pero
se utilizan con distintas nalidades. Se preere el DNA, ms
estable y con una doble hlice, para conservar y transmitir la
informacin gentica, y el RNA se deja que cumpla la funcin de dirigir la sntesis de protenas (RNA mensajero) y de
catlisis (RNA ribosomal).
En resumen, es muy probable que el RNA fuera el primero en la evolucin, con sus propiedades genticas y catalticas.
Una vez que la sntesis proteica se organiz, el DNA sustituy
al RNA en las funciones genticas, mientras las protenas se
ocupaban en las funciones catalticas. El RNA se mantuvo
como producto intermediario interconectando ambos. Con
toda seguridad, el DNA surgi cuando la creciente complejidad de la informacin gentica necesaria para la viabilidad
de la clula primitiva requiri una forma ms estable que la
que poda ofrecer el RNA.
sencillas que se supone surgieron hace menos de 3 500 millones (g. 1-13).
Las clulas procariotas son los organismos ms sencillos
que se encuentran prcticamente en todos los ambientes naturales. Tienen una forma esfrica o abastonada, y miden slo
algunos micrmetros. En ocasiones poseen una pared celular
protectora por fuera de la membrana celular que engloba un
nico compartimiento que contiene DNA, RNA, protenas
y otras pequeas molculas. A veces disponen de agelos y de
organelos aptos para la fotosntesis. Las fotografas obtenidas
con microscopio electrnico de los organismos procariticos
muestran una matriz de textura variable sin ninguna estructura
organizada.
Estos organismos se multiplican por divisin. Cuando
las condiciones ambientales son adecuadas (nutrimentos,
oxgeno, temperatura, etc.) una nica clula procariota se
puede dividir cada 20 minutos, con lo cual origina una poblacin de unos 5 000 millones de individuos en 11 horas. Esta
capacidad de multiplicacin es indicio de que las bacterias,
que son organismos unicelulares, se adaptan con rapidez al
ambiente. Por consiguiente, en condiciones de laboratorio,
se puede lograr que una bacteria experimente una mutacin
De la clula procariota
a la eucariota
Clulas procariotas
Un hito importante en la historia de la evolucin fue la aparicin de las primeras clulas procariotas, clulas relativamente
Figura 1-13.
Captulo 1 La clula: evolucin y desarrollo de los sistemas celulares
espontnea y evolucione para adaptarse a un nuevo azcar
como fuente de carbohidratos, o bien, que se haga resistente
a determinado antibitico.
Las bacterias se clasican en dos grandes grupos de
acuerdo con su hbitat (g. 1-14):
las eubacterias, que viven en los ambientes normales
(suelo, agua, plantas y otros organismos superiores)
las archaebacterias, que se han adaptado a vivir en condiciones sumamente adversas (fondo del mar, fuentes termales, zonas ricas en gases sulfurosos, etc.)
A pesar de su relativa simplicidad (o quizs por ello), las
bacterias se alimentan con todo tipo de sustancias: azcares
sencillos o complejos, grasas, protenas, hidrocarburos, polisacridos, etc. Algunas incluso pueden aprovechar el CO2
como fuente de carbono y el N2 como fuente de nitrgeno.
En la actualidad son, sin duda alguna, los habitantes ms
abundantes del planeta Tierra, han sobrevivido durante ms
tiempo y probablemente son los que sobrevivirn ms.
Reacciones metablicas
Una bacteria que crece en un medio salino glucosado y que es
su nica fuente de carbono lleva a cabo un enorme nmero de
reacciones qumicas porque utiliza la glucosa como fuente de
energa, pero tambin construye a partir de este azcar todas
las sustancias qumicas que necesita para su desarrollo y mul-
13
tiplicacin. Todas estas reacciones qumicas estn vinculadas
entre s a lo largo de la llamada cadena metablica.
En la clula primitiva, esta cadena metablica no tena
excesiva importancia, ya que la clula extraa las pocas molculas necesarias para su subsistencia de la sopa prebitica. Sin
embargo, al agotarse estos recursos naturales, los organismos
que tenan ventaja eran los que haban perfeccionado sistemas
enzimticos capaces de crear las molculas necesarias. Por
eso existe la opinin de que la cadena metablica aument
en complejidad hasta llegar a la presente en los organismos
actuales.
Si se plantea la hiptesis de que la cadena metablica
evolucion mediante sucesivos aadidos de pasos enzimticos,
est claro que las reacciones ms antiguas deben estar ms
prximas al centro del tronco metablico, que es donde se
sitan las reacciones ms vitales, y, adems, todas las clulas
vivientes las deben compartir. Este tronco metablico est
ocupado por la gluclisis, proceso mediante el cual la glucosa
se degrada en forma anaerobia (en ausencia de oxgeno, dado
que la atmsfera primitiva no contena oxgeno). La gluclisis
se verica virtualmente en todas las clulas y produce ATP
(trifosfato de adenosina), la fuente ms verstil de energa que
utilizan todas las clulas.
Junto a estas reacciones de los fosfatos de azcares se
ejecutan cientos de otras reacciones: mediante algunas se
sintetizan pequeas molculas que luego son utilizadas para
generar molculas ms complejas, otras son empleadas para
degradar grandes molculas, que llegan por medio de los
alimentos, en molculas ms pequeas. Estas reacciones
bacterias anaerbicas capaces
de vivir a altas temperaturas
(p.ej., bacterias sulfurosas)
ARCHAEBACTERIA
bacterias capaces de vivir con
altas concentraciones de sal
(bacterias halfilas)
bacterias que reducen el
metano a CO2 (bacterias
metangenas)
PROCARIOTA
ANCESTRAL
bacterias grampositivas
bacterias verdes fotosintticas
cianobacterias (algas azul-verdosas)
EUBACTERIA
bacterias prpura fotosintticas
bacterias gramnegativas
espiroquetas
Figura 1-14. Evolucin de las clulas procariotas.
14
Captulo 1
La clula: evolucin y desarrollo de los sistemas celulares
metablicas son comunes a todos los seres vivos. Evidentemente, existen diferencias: algunas especies no son capaces
de producir determinadas sustancias (p. ej., el hombre y otros
animales superiores no pueden fabricar el aminocido lisina,
que sintetiza cualquier bacteria), pero en general existe una
amplia concordancia de los procesos qumicos de todos los
seres vivos, prueba de una evolucin a partir de una clula
primitiva comn.
Fotosntesis
Si los primeros procesos metablicos evolucionaron para llenar
el vaco de compuestos orgnicos de la sopa prebitica, cabe
preguntarse qu pas cuando se agotaron estos compuestos.
Est claro que los organismos capaces de utilizar como fuente
de carbono el CO2 y como fuente de nitrgeno el N2 atmosfrico tenan ms ventajas de sobrevivir. Sin embargo, aunque
son muy abundantes en la atmsfera, ambos compuestos son
muy estables, por lo cual se requiere mucha energa y un
nmero elevado de reacciones para convertirlos en productos
sencillos, como azcares o aminocidos.
En el caso de la captacin del CO2 se cre un mecanismo,
la fotosntesis, mediante el cual un pigmento, la clorola,
capta la energa radiante procedente del Sol. La clula utiliza
posteriormente esta energa para sintetizar las molculas que
necesita.
Una de las primeras reacciones que induce la luz solar
fue, quiz, la fosforilacin de los nucletidos para originar
el ATP, comodn energtico de las clulas. Otra reaccin
probable fue la reduccin del CO2 y del N2 para permitir su
incorporacin a molculas orgnicas. La reduccin incorporacin de electrones se efectu al inicio a partir del SH2 y
luz solar, y despus a partir de H2O y luz solar. As empez la
produccin en gran escala del O2, como producto secundario
de la fotosntesis. Las cianobacterias (tambin llamadas algas
azules) son microorganismos actuales capaces de jar el CO2 y
el N2 de la atmsfera, y quizs sean los organismos ms autosucientes que existan ya que, en principio, pueden vivir slo
de agua y luz solar. Junto con otros organismos que tienen estas
propiedades, las cianobacterias, que se mantienen constantes
desde hace miles de millones de ao, crearon las condiciones
idneas para que otros organismos pudieran evolucionar con
mayor facilidad.
Respiracin
Por ser el oxgeno una sustancia altamente reactiva, debi resultar txica para los organismos primitivos, de la misma manera
que actualmente es txica para los microorganismos anaerobios. Por esta razn y para aprovechar su energa qumica, las
clulas primitivas aprendieron a utilizar este elemento para
CH2OH
OH
OH
O
C
OH
OH
2ADP3
+2Pi2
2ADP4
d-Glucosa-
OH
CH3
l-Lactato
Figura 1-15. Esquema simplificado de la gluclisis.
mejorar los procesos de degradacin de las molculas alimenticias. La glucosa, en ausencia de oxgeno, se descompone en
cido lctico o etanol (y produce dos molculas de ATP), pero
en presencia de oxgeno se degrada a CO2 y H2O y produce 36
molculas de ATP. La clula utiliza la energa producida en
este proceso, denominado respiracin, para su metabolismo y
produccin de nuevas molculas de la misma manera que con
la fotosntesis se obtiene ATP en los organismos fotosintticos. La mayor parte de los organismos, incluso casi todos los
procariotas, emplean hoy el proceso de la respiracin.
Gluclisis
La ruta glucoltica (o va de EmbdenMeyerho) consiste en
la degradacin anaerbica de la glucosa a lactato. Para muchos
tejidos, la gluclisis representa una ruta metablica de urgencia, capaz de producir dos moles de ATP a partir de una mol
de glucosa en ausencia de oxgeno molecular (g. 1-15).
La clula eucariota
La clula eucariota tiene en la actualidad una estructura organizada. A medida que la atmsfera se enriqueca con oxgeno,
pereci una parte de las clulas primitivas que no pudieron
adaptarse a estas nuevas condiciones. Otras desarrollaron
una capacidad para respirar, o bien, tuvieron que ocultarse
en lugares donde el oxgeno estaba ausente para conservar
su condicin de anaerobias. Sin embargo, una tercera clase
descubri que si se una en simbiosis con una clula aerobia
poda sobrevivir y desarrollarse de un forma mucho ms rica.
sta es la hiptesis ms viable para la organizacin metablica
de las clulas eucariotas de hoy da que se estudia en detalle
ms adelante (cuadro 1-1).
Por denicin y en contraste con las clulas procariotas, las clulas eucariotas tienen una estructura organizada
y disponen de un cierto nmero de organelos. En particular,
tienen un ncleo, separado del resto de la clula mediante
Captulo 1 La clula: evolucin y desarrollo de los sistemas celulares
15
Cuadro 1-1. Teoras que explican la evolucin de las clulas eucariotas. (Gupta, et al. TIBS, 1996;2:166-171.
Teoras sobre el origen de la clula eucariota
Se propone el origen simbitico de la clula eucariota
Mereschkowsky
1905
Origen endosimbitico de las mitocondrias, cloroplastos, citoesqueleto y del ncleo a partir de
grupos especficos de bacterias.
Margulis
1970
Origen independiente de las archaebacteria, eubacteria y eucariotas a partir de un ancestro
universal.
Woese
1981
Suposicin de que el ncleo podra ser una especie de Archaebacteria.
Lake, et al.
1982
Se propone la evolucin de la clula eucariota a partir de una clula intermedia entre
archaebacteria y una bacteria gramnegativa. Es opuesta al origen endosimbitico del ncleo
Cavalier-Smith
1987
Evolucin directa del genoma nuclear de los eucariotas a partir de una archaebacteria ancestral.
Iwabe, et al
Gogarten, et al
Woese, et al
1989
1989
1990
La evolucin de la clula eucariota a partir de un linaje protoeucariota, que presentaba metabolismo
basado en RNA, que engull a las clulas procariotas (archaebacterias)
Hartman
Sogin
1984
1991
La posibilidad de que el genoma nuclear de las clulas eucariotas est formado qumicamente por
una fusin primaria de una archaebacteria y una eubacteria.
Zillig
1991
El grupo eocito de archaebacteria fue sealado como el grupo ms prximo a los eucariotas.
Rivera y Lake
1992
una membrana nuclear, que es casi por completo de DNA.
El resto de la clula est constituido por el citoplasma, lugar
donde se realiza la mayor parte de las reacciones metablicas
y donde se encuentra un cierto nmero de organelos. stos
se tratan con mayor detalle ms adelante; entre ellos destacan
(g. 1-16):
Vescula
Membrana
plasmtica
Lisosoma
Aparato
de Golgi
Microfilamento
Citosol
Peroxisoma
Retculo
endoplsmico
Mitocondria
Polisoma
Microtbulo
Ribosoma
Ncleo
Figura 1-16.
Membrana
nuclear
Flagelo
16
Captulo 1
La clula: evolucin y desarrollo de los sistemas celulares
mitocondrias y cloroplastos
retculo endoplsmico liso (no) y retculo endoplsmico
rugoso (grueso)
aparato de Golgi
ribosomas
lisosomas y peroxisomas
citoesqueleto
vacuolas
agelos
Una membrana ms o menos organizada rodea la clula eucariota, como las procariotas; dicha membrana est
constituida fundamentalmente por fosfolpidos y protenas
especcas que tienen diferentes funciones.
Al parecer, las mitocondrias evolucionaron a partir de algn
organismo procariota que se uni en simbiosis con algn otro
organismo primitivo anaerobio. En efecto, en muchos aspectos,
las mitocondrias se parecen a los organismos procariotas:
tienen un tamao y forma parecidos
se multiplican por divisin
contienen su propio DNA
el proceso de respiracin se efecta exclusivamente en
ellas
muchas bacterias actuales respiran igual que las mitocondrias
Mediante esta simbiosis, primitivos eucariotas anaerobios
podran haber sobrevivido en un ambiente cada vez ms rico
en oxgeno, utilizando la capacidad del procariota asociado
para producir energa mediante el consumo de oxgeno atmosfrico.
La adquisicin de mitocondrias tuvo, sin duda, importantes repercusiones. Al ocuparse slo de la respiracin,
permiti que la membrana se especializara en otras funciones,
a diferencia de lo que ocurre en los procariotas; en stos se
debe conservar un gradiente importante de H+ en la membrana
plasmtica para producir ATP. Por consiguiente, la membrana
Archaebacteria
Otras
eubacterias
Bacterias
prpuras
sufurosas
eucariota pudo crear canales inicos y otros organelos receptores especializados en el intercambio de seales.
Los cloroplastos llevan a cabo la fotosntesis de la misma
manera que las cianobacterias: absorben la luz solar con la clorola que existe en sus membranas. Tambin los cloroplastos
maniestan similitudes notables con las cianobacterias:
son similares en forma y tamao
las membranas en las que se encuentra la clorola estn
dispuestas en capas
se reproducen por divisin
contienen DNA con secuencias muy parecidas al DNA
bacteriano
Todo ello apunta a que los cloroplastos evolucionaron
a partir de las cianobacterias que se unieron en simbiosis a
alguna clula eucariota primitiva. Este tipo de simbiosis es
relativamente frecuente y, en la actualidad, se conocen clulas
eucariotas que tienen autnticas cianobacterias en su interior.
La teora de la evolucin de los eucariotas a partir de la hiptesis simbitica se ilustra en la gura 1-17.
Teora actual de la
endosimbiosis como origen
de la clula eucariota
La clula eucariota se distingue tambin por una red proteica, el citoesqueleto, que sita y desplaza los organelos en
la clula. Todos los vegetales, los hongos y los animales son
eucariotas. Su DNA est en uno de los organelos, el ncleo.
Adems del retculo endoplsmico y el complejo de Golgi,
que intervienen en la sntesis de ciertas protenas, los otros
dos compartimientos que desempean un papel sustancial
ya que suministran energa a la clula son las mitocondrias
Cianobacterias
Plantas
Cloroplastos
Mitocondrias
Eucariota
ancestral
Procariota ancestral
anaerobio
Procariota ancestral
Figura 1-17. Teora simbitica de la evolucin de los eucariotas.
Hongos Animales
Captulo 1 La clula: evolucin y desarrollo de los sistemas celulares
(del griego mitos, lamento, y khondros, grano), las cuales se
ocupan de la respiracin celular, y los plastos de los vegetales,
donde tiene lugar la fotosntesis o conversin de la energa
luminosa en energa qumica.
Algunos organelos pueden desaparecer y volverse a
formar. Es el caso del ncleo durante la divisin celular,
del aparato de Golgi y del retculo. Pero las mitocondrias y
los plastos proceden siempre de la divisin de mitocondrias
y plastos preexistentes. A nales del siglo XIX, esta particularidad haca pensar ya en bacterias que se dividan por
biparticin en el citoplasma. Las mitocondrias y los plastos
eran organismos vivos, instalados en simbiosis en la clula.
En 1883, el investigador alemn A. Schimper observ: Si se
establece denitivamente que los plastos nunca se forman de novo
en la clula huevo, entonces su situacin en la clula donde se encuentran recuerda la de los simbiontes. Es posible que una planta
verde no sea otra cosa que la unin entre un organismo incoloro y
un microbio que posee los pigmentos cloroflicos. Otros autores
le pisaron los talones a principios del siglo XX, pero toparon,
lamentablemente, con la incredulidad de sus colegas. Ocurri
como con la deriva de los continentes de Alfred Wegener: los
argumentos propuestos no bastaron para aanzar la intuicin.
En particular, fracasaron todos los intentos de aislamiento y
cultivo in vitro del microbio verde. Varios autores propusieron
para estos organelos un origen autgeno a partir de repliegues
de la membrana plasmtica. Los progresos de la biologa iban
a brindar otros argumentos a la idea segn la cual las mitocondrias y los plastos derivan de bacterias.
La microscopia electrnica demostr que los organelos
de los eucariotas estn limitados por una membrana lipdica
simple, salvo tres de ellos: el ncleo, rodeado por membranas
provistas de poros que lo ponen en continuidad con el citoplasma, y los plastos y las mitocondrias, separados del citoplasma
por dos membranas continuas. Adems, la organizacin de la
estructura de estos dos ltimos organelos evoca la de ciertas
bacterias. Por ejemplo, las membranas fotosintticas o tilacoides de los plastos de ciertas algas (las algas rojas) evocan en
gran medida a los tilacoides de bacterias fotosintticas, las
cianobacterias. Los adelantos de la bioqumica pusieron de
maniesto otros parecidos sorprendentes. Los lpidos de las
membranas que rodean a las mitocondrias existen en ciertas
bacterias, pero no en otras partes de la clula eucariota. En
cuanto a los lpidos caractersticos de las membranas de los
plastos, se les encuentra en las cianobacterias. Adems, las
mitocondrias y los plastos desempean ciertas funciones
metablicas que efectan tambin ciertos procariotas libres.
La fotosntesis se consuma de manera casi idntica en las cianobacterias y los plastos. Las reacciones de oxidorreduccin
ligadas a la actividad energtica de las mitocondrias y de los
plastos requieren la intervencin de cadenas de transferencia
de electrones membranosas parecidas a las de los procariotas:
colaboran en ellas unas complejas protenas homlogas, como
los citocromos.
17
Pero el argumento principal fue el descubrimiento de un
genoma en los plastos en 1962 y luego en las mitocondrias
al ao siguiente. Este DNA se parece al de los procariotas:
no est aislado dentro del organelo y se presenta en forma
de varias copias idnticas de una molcula circular. El DNA
se duplica y se transere durante la biparticin del organelo
como en el caso de los procariotas libres. En las mitocondrias
y los plastos existen ribosomas que sintetizan protenas y son
ms pequeos que los del citoplasma de la clula. De hecho,
tienen el tamao y la composicin del RNA de los ribosomas
de los procariotas.
Por todas estas razones, la hiptesis del origen simbitico
de las mitocondrias y de los plastos hall adeptos en los aos
1970, como Lynn Margulis, de la Universidad de Amherst
(Massachusetts), y Sarah P. Gibbs, de la Universidad McGill
en Montreal. La biologa molecular subray todava ms el
parentesco entre estos organelos y los procariotas. Es el caso,
por ejemplo, de las zonas promotoras de los genes, es decir,
de las secuencias reconocidas para protenas que aseguran su
transcripcin en RNA. Se descubrieron actualmente genes
homlogos (que tienen el mismo origen evolutivo) de genes
bacterianos en el genoma de las mitocondrias y de los plastos. Sera sorprendente encontrar bacterias en el citoplasma
de eucariotas. En realidad, semejante modo de vida no es
raro entre los eucariotas: se le designa endosimbiosis. Este
modo de vida es el de ciertos microorganismos patgenos,
como la bacteria causante de la listerosis o de la ebre tifoidea. Pero tambin, el de numerosos animales unicelulares,
los protozoarios, que son colonizados por varias bacterias.
Kwang W. Jeon, de la Universidad de Tennessee, describi
la aparicin de una endosimbiosis entre las amebas, a raz de
una infeccin que diezm un cultivo de laboratorio en 1966.
Algunas amebas sobrevivieron a la presencia de bacterias en
su citoplasma: veinte aos ms tarde, todava contienen unas
42 000 bacterias por clula. Pero hay ms: esta asociacin se
ha vuelto forzosa. Las amebas ya no pueden sobrevivir si se
da muerte a sus compaeros citoplasmticos aumentando la
temperatura. Otro ejemplo clsico: las plantas de la familia
de las leguminosas (trbol, guisante) poseen en sus races
unos ndulos cuyas clulas albergan bacterias del gnero
Rhizobium. Estas bacterias, llegadas del suelo, ayudan a la
planta a alimentarse jando el nitrgeno atmosfrico. Estos
microorganismos unicelulares estn rodeados siempre por dos
membranas: la membrana interna, que es la de la bacteria, y la
membrana externa procedente de la membrana plasmtica del
hospedero, que rodea a la bacteria durante su penetracin en
la clula. No obstante, el origen de la membrana externa de
las mitocondrias y los plastos es objeto de controversia (en el
caso de los plastos, contiene lpidos de tipo procariota y sera
de origen mixto).
Cuntos sucesos de endosimbiosis han tenido lugar en
el curso de la evolucin? Cules son los procariotas actuales
ms prximos a los que viven en endosimbiosis? La biologa
18
Captulo 1
La clula: evolucin y desarrollo de los sistemas celulares
molecular actual puede empezar a responder estas preguntas
gracias a la comparacin de los genes de los organelos con
los genes homlogos de los procariotas. Las secuencias de
los genes se alinean y luego se comparan visualmente o por
medio de programas de computadora que descubren parentescos en forma de rboles logenticos. Se averigua as que
las mitocondrias forman un grupo homogneo: todas ellas
descenderan de una bacteria ancestral nica. Se supone que
un solo suceso de endosimbiosis se produjo en un antepasado
de la mayora de los eucariotas actuales.
Se conocen descendientes directos de estos eucariotas
primitivos, como el parsito intestinal Giardia. Trabajos
recientes indican que el ancestro de las mitocondrias es
parecido a un grupo de bacterias que usa oxgeno y que engloba, entre otros, al agente de la ebre tifoidea, Rickettsia, y
a varias bacterias simbiticas de artrpodos. Las bacterias de
este grupo comparten varios caracteres con las mitocondrias,
como la vida intracelular obligatoria, la utilizacin de oxgeno
y la produccin de una pequea molcula de transferencia de
energa, el ATP (trifosfato de adenosina), por medio de una
va metablica llamada ciclo de Krebs.
Tal vez, las mitocondrias se consolidaron hace 2 000 o 3
000 millones de aos, en el periodo precmbrico, lo cual constituye una etapa fundamental para los eucariotas. Esta etapa
supone la ventaja de la respiracin celular: sin mitocondrias,
la clula eucariota no puede utilizar el oxgeno para oxidar los
azcares y producir su energa. Es posible incluso que los eucariotas primitivos fueran sensibles al oxgeno, como lo son en la
actualidad los eucariotas carentes de mitocondrias. Las primeras
mitocondrias, al consumir el oxgeno, pudieron desempear el
papel de protector ante las clulas que las albergaban: el oxgeno
genera derivados txicos para las clulas que no poseen las
enzimas necesarias para inactivarlos.
En la actualidad, son raros los eucariotas desprovistos
de estos organelos: dicha endosimbiosis parece haber sido
determinante para el xito evolutivo de los eucariotas.
Ms tarde, ciertos eucariotas que originaron los diversos
linajes de vegetales habran logrado conformar los plastos,
tal vez hace alrededor de 1 200 o 2 000 millones de aos. La
diferencia entre estos linajes son los pigmentos fotosintticos
que utilizan para captar la energa luminosa. Ya se mencionaron las algas rojas. Su equipo pigmentario recuerda el de las
cianobacterias (clorola a y cobilisomas que les dan su color
rojo). Se distinguen tambin los vegetales verdes, que agrupan
las algas verdes y sus descendientes, los vegetales terrestres:
carecen de pigmentos proteicos rojos, pero estn dotados de
clorola tipo a y b. La informacin molecular indica que los
plastos verdes y rojos habran surgido a partir de un mismo
linaje. Las cianobacterias actuales son, a menudo, simbiticas y
se unen a hongos, helechos o, incluso, viven en endosimbiosis
en protozoos o esponjas, por ejemplo.
Lo que puede sorprender, en cambio, es que el equipo
pigmentario de los plastos verdes es muy distinto al de las
cianobacterias. La prdida de los cobilisomas y la produccin
de la clorola b se produjeron varias veces en las cianobacterias
durante su evolucin. En 1975, Ralph A. Lewin, de la Universidad de Yale, descubri Prochloron, un procariota carente
de cobilisomas pero que posee clorola b. Desde entonces se
han descubierto otros organismos parecidos y se ha propuesto
crear para ellos el grupo de los proclorotos. Pero la informacin
molecular indica que, en realidad, estos organismos aparecieron en diversas ocasiones entre las cianobacterias y forman
linajes diferentes, sin ninguna relacin con los plastos de los
vegetales verdes. Si bien los ancestros de los plastos rojos y
verdes son prximos, por no decir idnticos, parece probable
que en el origen de las algas rojas y las plantas verdes hubiera
dos hospederos eucariotas diferentes.
En ciertas algas, los plastos presentan la impensada particularidad de estar rodeados no de dos, sino de cuatro membranas. Cmo explicarlo? Estas algas pertenecen al grupo
de las heterocontas: comprende las algas pardas de las costas
mexicanas, como el fucus, pero tambin las microscpicas diatomeas. Su plasto pardo, distinto de los plastos verdes y rojos,
posee las clorolas de tipos a y c. Entre la segunda y tercera
membranas, un espacio citoplasmtico contiene vesculas y
microtbulos. Cmo aparecieron estos plastos especiales? Un
pequeo grupo de algas unicelulares, las criptotas, atrajo la
atencin: estas algas tambin estn provistas de plastos rodeados de cuatro membranas. Pero en el espacio entre la segunda
y la tercera membrana hay ribosomas de tipo eucariota y un
organelo curioso, el nucleomorfo, que evoca un ncleo reducido que contiene DNA. Sarah P. Gibbs plante la hiptesis
de una segunda endosimbiosis para explicar estos plastos de
cuatro membranas. Entonces, el plasto comprende, del exterior al interior, una membrana de secuestro del hospedero
secundario, la membrana plasmtica de un alga asimilada y
luego las dos membranas del plasto de esta ltima.
De hecho, las algas unicelulares tienen la capacidad de
desarrollarse perfectamente en el citoplasma de otros eucariotas, por ejemplo, en las clulas de los animales que edican
los corales y las anmonas de mar de las costas mexicanas. Las
algas en cuestin, ciertas dinoageladas, existen tambin en
forma libre y son todava reconocibles a la perfeccin. Tales
estados intermedios pudieron originar asociaciones ms estrechas, como en las criptotas y en las algas heterocontas.
El DNA del nucleomorfo de las criptotas contiene genes
que corresponden a los RNA de los ribosomas que lo rodean.
El genoma del nucleomorfo est formado por tres pequeos
cromosomas. Estos genes son parecidos a los de las algas rojas,
y los genes ribosmicos del ncleo de la clula hospedera,
la criptota, hacen que sta est ms emparentada con los
vegetales verdes (g. 1-18).
Las algas heterocontas, por su parte, tambin habran
integrado un alga roja, la cual habra sufrido una regresin
mayor. Los trabajos acerca del genoma procaritico contenido
en el seno de los plastos pardos muestran la liacin de ste
Captulo 1 La clula: evolucin y desarrollo de los sistemas celulares
19
Mitocondrias de animales
Mitocondrias
de hongos Vaca Erizo
Cachliabalus
Padospora
Aspergillus
Acanthamoeba
(protozoos)
Pylalello
(alga parda)
Mitocondrias de
plantas
Maz Tabaco
Anagra
Marchantia
(hopatica)
Prototheca
(alga verde)
Chondrus
(alga roja)
Mitocondrias
Rhodospirillum
rubrum
Protobacterias
Synechocystis
Anabaena
Cianobacterias
Figura 1-18.
con respecto a los plastos de algas rojas. En el transcurso de la
evolucin, el alga roja asimilada habra perdido sus pigmentos
rojos y adquirido la clorola c, con lo cual se convirti en un
plasto pardo caracterstico.
Otros organismos pudieron adquirir plastos por endosimbiosis secundaria. Esto resuelve la vieja paradoja de las
Euglena. Los manuales presentan a estos protozoarios como
mitad animales (ya que las Euglena son agelados y pueden
fagocitar presas o restos) y mitad vegetales (pues poseen
plastos con los mismos pigmentos que los de los vegetales
verdes). En realidad, como se demuestra al comparar los genes
de los plastos (g. 1-19), se trata efectivamente de plastos
de vegetales verdes. No obstante, las Euglena son eucariotas
primitivos muy cercanas a los tripanosomas. Pudieron adquirir
sus plastos a partir de un alga verde, presa o endosimbionte.
Numerosos ejemplos actuales ponen de maniesto fases
intermedias en las endosimbiosis, tanto primarias como secundarias. De hecho, el aprendizaje de la vida en comn tiene
numerosas exigencias. El encuentro de los participantes es la
primera fase, que no necesariamente representa ventaja para
los dos. Las mitocondrias derivan, tal vez, de bacterias patgenas endosimbiticas, como las bacterias actuales causantes
de la listerosis o de la ebre tifoidea. Por lo que se reere a
las cianobacterias, fueron tal vez presas antes de convertirse
en plastos. En una segunda fase, el vnculo puede volverse
ms duradero. Puede signicar una ventaja para un eucariota
cazador de cianobacterias aprovechar algn tiempo la fotosntesis de su presa antes de digerirla, o bien, criarla en su
citoplasma. Cuando la relacin es permanente, es necesario
un control recproco sobre la multiplicacin de cada socio
Escherichia
coli
Maz
Antilharmnion
Guisante
(alga roja)
Euglena Marchantia
Odontella
(hepatica)
(diatomea)
Plastos
Figura 1-19.
para la supervivencia del organismo quimera formado. Queda
abierto entonces el camino para una evolucin conjunta (o
coevolucin) de los socios, y pueden establecerse intercambios
recprocos.
Para el endosimbionte, la vida en el interior de la clula
es en extremo confortable; es el hospedero el que soporta las
agresiones del medio externo. Este hbitat protegido explica
la desaparicin de las envolturas protectoras; en los plastos
con dos membranas ya no quedan huellas de la pared de las
cianobacterias. La regresin de la pared tambin ha sido
descrita en algas eucariotas unicelulares endosimbiticas actuales. Pero la evolucin regresiva ms curiosa es, en verdad,
la reduccin del genoma de los organelos con respecto a las
formas libres en el transcurso de la evolucin (vase cuadro
1-2). Ya desapareci el ncleo del alga roja que asimilaron las
algas heterocontas de plastos pardos; el tamao del genoma
de tipo procariota que est contenido en los organelos es entre
diez y cien veces ms pequeo que el de los procariotas libres.
Por supuesto, ya desaparecieron todos los genes necesarios
para orientarse, desplazarse y protegerse en el medio exterior,
20
Captulo 1
La clula: evolucin y desarrollo de los sistemas celulares
pero otros siguen existiendo: simplemente fueron transferidos
al ncleo de la clula hospedera mediante un proceso que se
empieza a entrever gracias al estudio reciente de Brennicke y
colaboradores (cuadro 1-2).
En el curso de la evolucin, muchos genes procedentes
de los plastos y las mitocondrias migraron hacia el ncleo de
la clula que los albergaba; aunque son nucleares, estos genes
tienen, entonces, un origen evolutivo procariota.
Estas transferencias sellan la asociacin y son la razn de
que no se puedan cultivar de modo aislado los organelos, los
cuales se volvieron por completo dependientes de su hospedero.
La simbiosis se volvi forzosa y los socios forman un nuevo organismo quimrico. Unas regulaciones acoplan la expresin de
ambos genomas, y participan en el buen funcionamiento de la
nueva unidad siolgica. La clula eucariota esconde tal vez otras
endosimbiosis. Es el caso de los peroxisomas, unos organelos que
intervienen en ciertas reacciones metablicas como la fotorrespiracin y las peroxidaciones. Estos organelos estn limitados por
una simple membrana, carecen de DNA y aparecen siempre por
biparticin. Algunos cientcos los consideran como endosimbiontes cuyo genoma fue transferido del todo al ncleo.
Por tanto, la endosimbiosis inuy profundamente en la
evolucin de los eucariotas y les facilit la consecucin de las
potencialidades metablicas de los procariotas. Este fenmeno
acompa varias veces la fundacin de linajes evolutivos en
los eucariotas (g. 1-20). Se complementan las potencialidades morfogenticas de los eucariotas (citoesqueleto, paso
al estado pluricelular, entre otros) y la ingeniera enzimtica
(jacin del carbono o del nitrgeno, respiracin, etc.) de los
procariotas, los cuales, aunque a menudo asimilados, nunca
albergan simbiontes.
Cuadro 1-2. Reduccin del endosimbionte
Organismo
Nmero
total
de genes
Tamao del genoma
(en pares de bases)
Cianobacterias libres
> 5 000
de 1 a 10 millones
Porphyra Alga roja
210
183 000
Cryptomonas
Criptofita
140
112 000
Oryza (arroz)
110
114 000
Nicotiana (tabaco)
113
131 000
Tamao de los genomas procariotas contenidos en los
plastos de distintos grupos de vegetales comparado con
las cianobacterias libres. En el transcurso de la evolucin,
numerosos genes de los plastos se transfirieron al ncleo del
hospedero eucariota.
Adaptado de Douglas SE, et al. Nature, 1991;350:148.
plantas
terrrestres
algas verdes
clorobiontes
rizpodos
criptfitas
(ncleo)
oomicetos
(seudohongos)
diatomeas
y algas pardas
protistos
criptfitas
(nucleomorfo)
heterocontas
rodobiontes
algas rojas
esporozoos
(plasmodium)
dinoflagelados
alveolobiontes
ciliados
(paramecios)
distintos grupos
de protozoos (amebas)
tripanosomas
Figura 1-20. Antepasados comunes con los procariotas.
Los sucesos de endosimbiosis parecen haber sido bastante numerosos, en ocasiones sucesivos, y originaron los
plastos de cuatro membranas. En la actualidad, ciertas algas
unicelulares, las criptotas y las heterocontas, cuyo plasto de
cuatro membranas deriva, recurdese, de una endosimbiosis
secundaria, viven en simbiosis en el citoplasma de dinoagelados que perdieron sus propios plastos. Se trata entonces de
tres endosimbiosis sucesivas.
Estructura celular
Las clulas son estructuras con muy buena organizacin en
su interior; estn constituidas por diferentes organelos, y cada
uno de ellos cumple funciones diferentes. Sin embargo, todas
las clulas eucariotas, que son las de todos los seres vivos con
excepcin de las bacterias, cuyas clulas son mucho ms sencillas, comparten un plan general de organizacin:
Una membrana que determina su individualidad
Un ncleo que contiene el material gentico y ejerce el
control de la clula
Captulo 1 La clula: evolucin y desarrollo de los sistemas celulares
Un citoplasma lleno de organelos, dnde se ejecutan
prcticamente todas las funciones.
La gura 1-21 es un dibujo esquemtico de una clula
eucariota animal y de otra vegetal.
La clula est rodeada por una membrana, denominada
membrana plasmtica. La membrana delimita el territorio
de la clula y controla el contenido qumico de la misma.
Lpidos, protenas y glcidos forman parte de la composicin
qumica de la membrana, en proporciones aproximadas de 40,
50 y 10%, respectivamente, de acuerdo con la clula u organelo
que cubra. En la membrana interna mitocondrial hay hasta
85% de protenas, y en el sistema nervioso, slo 15%. El tipo
de lpidos tambin vara entre clulas. Los lpidos forman una
capa doble y las protenas se disponen de una manera irregular y asimtrica entre ellos. Estos componentes son mviles,
lo que conere a la membrana un elevado grado de uidez.
Por el aspecto y comportamiento el modelo de membrana se
denomina modelo de mosaico uido.
Las funciones de la membrana se resumen en:
transporte, que es el intercambio de materia entre el
interior de la clula y su ambiente externo.
reconocimiento y comunicacin, gracias a molculas
situadas en la parte externa de la membrana, las cuales
actan como receptores de sustancias.
y mantener su medio interno estable. La membrana maniesta una permeabilidad selectiva, ya que permite el paso de
pequeas molculas, siempre que sean liplas, pero regula
el paso de molculas no liplas.
El paso a travs de la membrana posee dos modalidades,
una pasiva, sin gasto de energa, y otra activa, con consumo
de energa.
Transporte pasivo
Es un proceso de difusin de sustancias a travs de la membrana. Se produce siempre a favor del gradiente, es decir, del
ambiente donde hay ms hacia el medio donde hay menos.
Este transporte se efecta por difusin simple.
Difusin simple
Es el paso de pequeas molculas a favor del gradiente; se
puede producir a travs de la bicapa lipdica o a travs de
canales proteicos.
1.
Transporte a travs de membranas
La bicapa lipdica de la membrana acta como una barrera
que separa dos medios acuosos: el medio donde vive la clula
y el medio interno celular.
Las clulas requieren nutrimentos del exterior, deben
eliminar sustancias de desecho procedentes del metabolismo
2.
Difusin simple a travs de la bicapa. De este modo entran molculas lipdicas como las hormonas esteroides,
anestsicos como el ter y frmacos liposolubles, as como
sustancias apolares como el oxgeno y el nitrgeno atmosfrico. Tambin algunas molculas polares muy pequeas,
como el agua, el CO2, el etanol y el glicerol, atraviesan
la membrana por difusin simple. La difusin del agua
recibe el nombre de smosis.
Difusin simple a travs de canales. Se realiza mediante
las denominadas protenas de canal. As entran iones
como el Na+, K+, Ca2+, Cl. Las protenas de canal son
protenas que forman un oricio o canal interno, cuya
apertura est regulada, por ejemplo por ligandos, como
Ncleo
Membrana
Citoplasma
Animal
Figura 1-21. Clula eucariota.
21
Vegetal
22
3.
Captulo 1
La clula: evolucin y desarrollo de los sistemas celulares
ocurre con los neurotransmisores o las hormonas, que se
unen a una determinada regin, o el receptor de la protena de canal, que sufre una transformacin estructural
que induce la apertura del canal.
Difusin facilitada. Permite el transporte de pequeas
molculas polares, como los aminocidos, monosacridos,
etc., que al no poder atravesar la bicapa lipdica, requieren
que protenas transmembrana faciliten su paso. Estas protenas reciben el nombre de protenas transportadoras o
permeasas, las cuales, al unirse a la molcula que tienen
que transportar sufren un cambio tal en su estructura que
arrastra a dicha molcula hacia el interior de la clula.
Transporte activo
citosol por su aspecto uido. En esta parte se encuentran las
molculas necesarias para el mantenimiento celular.
El citoesqueleto consiste en una serie de bras que da
forma a la clula, y conecta distintas partes celulares, como si
se tratara de vas de comunicacin celulares. Es una estructura
en cambio continuo que est formada por los tres tipos de
componentes siguientes.
Microtbulos
Son lamentos largos, formados por la protena tubulina. Son
las piezas ms importantes del citoesqueleto y pueden formar
asociaciones estables, como los
En este proceso tambin actan protenas de membrana, pero
stas requieren energa, en forma de ATP, para transportar las
molculas al otro lado de la membrana. Tienen lugar cuando
el transporte se realiza en contra del gradiente electroqumico.
Son ejemplos de transporte activo la bomba de Na+/K+ y la
bomba de Ca2+.
La bomba de Na+/K+ requiere una protena transmembrana
que expele Na+ hacia el exterior de la membrana y conduce K+
hacia el interior. Esta protena acta contra el gradiente gracias
a su actividad como ATPasa, ya que rompe el ATP para
obtener la energa necesaria para el transporte. Mediante este
mecanismo, se envan tres Na+ hacia el exterior y se incorporan dos K+ en el interior, con la hidrlisis acoplada de ATP.
El transporte activo de Na+ y K+ tiene una gran importancia
siolgica. De hecho, todas las clulas animales gastan ms
del 30% del ATP que producen (y las clulas nerviosas ms
del 70%) para movilizar estos iones.
Citosol y citoesqueleto
Toda la porcin citoplasmtica que carece de estructura y
constituye la parte lquida del citoplasma recibe el nombre de
Figura 1-22.
centriolos, que son dos pequeos cilindros situados en el
interior del centrosoma (g. 1-22), exclusivos de clulas
animales. Con el microscopio electrnico se observa que
la parte externa de los centriolos est formada por nueve
tripletes de microtbulos. Los centriolos se cruzan formando un ngulo de 90
cilios y agelos, que son delgadas prolongaciones celulares
mviles que poseen bsicamente la misma estructura; la
diferencia entre ellas es que los cilios son muchos y cortos,
y los agelos son pocos y ms largos (g. 1-23). Constan
de dos partes: una externa que sobresale de la supercie de la
clula, est revestida con membrana plasmtica y contiene
un esqueleto interno de microtbulos llamado axonema,
y otra interna, que se denomina cuerpo basal, de la que
salen las races ciliares que coordinan el movimiento
Microfilamentos
Se sitan sobre todo en la periferia celular, bajo la membrana
y estn formados por hebras de la protena actina, trenzadas
en hlice, cuya estabilidad se debe a la presencia de ATP e
iones de calcio. Se ocupan de la contraccin muscular cuando
se ligan a los lamentos de miosina.
Captulo 1 La clula: evolucin y desarrollo de los sistemas celulares
Flagelo
23
Membrana
plasmtica
Ncleo
Retculo
endoplsmico
Cromosomas
Ribosomas
Microtbulos
Ribosomas
Mitocondria
Mitocondria
Retculo
endoplsmico
Microfilamentos
y filamentos
intermedios
Aparato
de Golgi
Microtbulo
Figura 1-24.
Centriolos
Cilios
Clula compuesta
Figura 1-23.
Filamentos intermedios
Estn formados por diversos tipos de protenas. Son polmeros
muy estables y resistentes, y abundan especialmente en el citoplasma de las clulas sometidas a fuertes tensiones mecnicas
(queratina, desmina) ya que su funcin consiste en distribuir las
tensiones, que de otro modo romperan la clula (g. 1-24).
Distribucin de los filamentos
del citoesqueleto en el citoplasma
Como se puede apreciar en la gura 24, los microtbulos
irradian desde una regin del citoplasma denominada centro
organizador de microtbulos o centrosoma.
Los microlamentos se encuentran dispersos en todo el
citoplasma, pero se concentran fundamentalmente abajo de
la membrana plasmtica.
Los lamentos intermedios se extienden por todo el
citoplasma y se anclan a la membrana plasmtica, lo cual
proporciona resistencia mecnica a las clulas.
Retculo endoplsmico
Lo forman una red de membranas que originan cisternas,
sculos y tubos aplanados (g. 1-25). Delimita un espacio interno llamado lumen del retculo, y es una estructura continua
con la membrana externa de la envoltura nuclear.
Se pueden distinguir dos tipos de retculo, que son los
siguientes.
1.
2.
El retculo endoplsmico rugoso (RER) contiene ribosomas unidos a su membrana. En l se efecta la sntesis
proteica. Las protenas que sintetizan los ribosomas pasan
al lumen del retculo y ah maduran hasta ser exportadas
a su destino denitivo.
El retculo endoplsmico liso (REL) carece de ribosomas
y est formado por tbulos ramicados y pequeas vesculas esfricas. En este retculo se realiza la sntesis de
lpidos. En el retculo de las clulas del hgado tiene lugar
la desintoxicacin, que consiste en convertir una sustancia
o metabolito insoluble en agua en soluble en agua para
eliminar as dichas sustancias por la orina.
Aparato o complejo de Golgi
C. Golgi lo descubri en 1898. Consiste en un conjunto de
estructuras de membrana que forma parte del complicado
sistema de membranas, que se encuentra en el interior de las
clulas (g. 1-25). Es tanto ms complejo cuanto mayor es
la actividad celular.
La unidad bsica del organelo es el sculo, que consiste en
una vescula o cisterna aplanada. Cuando se apila una serie de
sculos, se forma un dictiosoma. Adems, es posible observar
toda una serie de vesculas ms o menos esfricas en ambos
lados y entre los sculos. El conjunto de todos los dictiosomas
y vesculas constituye el aparato de Golgi.
El dictiosoma se encuentra en ntima relacin con el retculo endoplsmico, lo que permite diferenciar dos caras: la cara
cis, ms prxima al retculo, y la cara trans, ms alejada. En la
cara cis se encuentran las vesculas de transicin, mientras que
en la cara trans, estn situadas las vesculas de secrecin.
24
Captulo 1
La clula: evolucin y desarrollo de los sistemas celulares
Aparato de Golgi
Vacuola
Vescula
Membrana
plasmtica
Lisosoma
Lisosoma
Aparato
de Golgi
Microfilamento
Citosol
Peroxisoma
Retculo
endoplsmico
Mitocondria
Polisoma
Microtbulo
Ribosoma
Ncleo
Membrana
nuclear
Flagelo
Peroxisoma
Retculo endoplsmico
liso
Retculo endoplsmico
rugoso
Figura 1-25.
El sistema de membranas constituye la respuesta de las
clulas eucariotas a la necesidad de regular sus comunicaciones
con el ambiente en el trasiego de macromolculas. Para ello
se desarrollaron dos mecanismos con los que el aparato de
Golgi guarda relacin.
La incorporacin de sustancias se lleva a cabo por endocitosis, mecanismo que consiste en englobar sustancias con
ayuda de la membrana plasmtica para asimilarlas posteriormente. La eliminacin de sustancias se efecta por exocitosis, mecanismo que consiste en la fusin de las vesculas que
contienen la sustancia por exportar con la membrana celular.
Estos mecanismos dan sentido funcional al aparato de Golgi
porque intervienen en:
la maduracin de las glucoprotenas provenientes del
retculo
los procesos de secrecin, almacenamiento, transporte y
transferencia de glucoprotenas
la formacin de membranas: plasmtica, del retculo,
nuclear
la generacin de la pared celular vegetal
la formacin de los lisosomas
Lisosomas
Los lisosomas tienen una estructura muy sencilla. Son semejantes a las vacuolas, los rodea slo una membrana y contienen
Captulo 1 La clula: evolucin y desarrollo de los sistemas celulares
25
gran cantidad de enzimas digestivas que degradan todas las
molculas inservibles para la clula (g. 1-25). Se forman a
partir del retculo endoplsmico rugoso. Posteriormente, el
complejo de Golgi empaqueta las enzimas.
Estos cuerpos funcionan como estmagos de la clula
o vacuolas digestivas. Adems de digerir cualquier sustancia
que ingrese del exterior, consumen restos celulares viejos para
digerirlos tambin, por lo que tambin se les llama vacuolas
autofgicas. Reciben, asimismo, el nombre de bolsas suicidas
porque si se rompiera su membrana, las enzimas encerradas
en su interior terminaran por destruir toda la clula.
En la membrana interna estn los sistemas dedicados al
transporte de los electrones que se desprenden en las oxidaciones anteriores y un conjunto de protenas encargadas de
acoplar la energa liberada del transporte electrnico con la
sntesis de ATP. Estas protenas le dan un aspecto granuloso
a la cara interna de la membrana mitocondrial.
Tambin se encuentras dispersas en la matriz unas
molculas de DNA circular y unos pequeos ribosomas que
sintetizan un pequeo nmero de protenas mitocondriales.
Mitocondria
Los cloroplastos son organelos exclusivos de las clulas vegetales (g. 1-26). En ellos tiene lugar la fotosntesis, proceso
en el que se transforma la energa lumnica en energa qumica
y se almacena en molculas de ATP y molculas reductoras
(NADPH + H+). Esta energa se utiliza despus para sintetizar
molculas orgnicas.
Tienen una organizacin muy similar a la de la mitocondria, aunque son de mayor tamao y poseen un compartimiento ms porque presentan un tercer tipo de membrana.
Por lo tanto, un cloroplasto contiene tres membranas y
tres compartimientos.
Cloroplasto
Las mitocondrias (g. 1-26) son los organelos celulares encargados de suministrar la mayor parte de la energa necesaria para
la actividad celular. Actan, por tanto, como centrales energticas de la clula y sintetizan ATP a expensas de los carburantes
metablicos (glucosa, cidos grasos y aminocidos).
La ultraestructura mitocondrial depende de las funciones
que desempea: en la matriz se localizan las enzimas que se
ocupan de oxidar cidos grasos, aminocidos, cido pirvico
y ciclo de Krebs.
Cloroplasto
Vescula
Membrana
plasmtica
Membrana
citoplsmica
Lisosoma
Aparato
de Golgi
Microfilamento
Citosol
Peroxisoma
Retculo
endoplsmico
Mitocondria
Polisoma
Microtbulo
Ribosoma
Ncleo
Ncleo
Figura 1-26.
Membrana
nuclear
Flagelo
Mitocondria
26
Captulo 1
La clula: evolucin y desarrollo de los sistemas celulares
La membrana externa es muy permeable, gracias a la
presencia de porinas.
La membrana interna es menos permeable y no presenta
pliegues (la de la mitocondria s los presenta). Entre ambas membranas queda un primer compartimiento, que
es el espacio intermembranas. La membrana interna delimita un espacio que es el estroma, donde hay ribosomas,
copias de DNA, distintos tipos de RNA, grnulos de
almidn y gotas de lpidos.
La membrana tilacoidal es el tercer tipo de membrana.
Forma unos sacos aplanados denominados tilacoides, y da
lugar a unas agrupaciones llamadas grana. Los tilacoides
estn interconectados y delimitan una tercera cavidad que
es el espacio tilacoidal. Esta membrana es la encargada de
captar la energa solar, gracias a la presencia de clorolas y
de otros pigmentos unidos a protenas en unas estructuras
funcionales que son los fotosistemas.
En esta membrana (g. 1-27) se encuentra tambin una
cadena de transporte de electrones y una sintasa de ATP, que
funciona como la sintasa mitocondrial de ATP.
En la gura 1-28 se ilustra la distribucin de estas estructuras en la membrana tilacoidal. El fotosistema II (PSII)
se localiza particularmente en la grana, mientras que el fotosistema I (PSI) est en contacto con el estroma al igual que
el complejo sintasa de ATP. El citocromo b6f tiene como
funcin el transporte de los electrones desde el fotosistema
II al I, por lo que se encuentra en ambos sitios.
un centro reactivo, o centro de reaccin fotoqumico, que
posee unas molculas de clorola a que actan como una
verdadera trampa energtica, puesto que los electrones
que liberan son enviados hacia la cadena de transporte
de electrones de la membrana tilacoidal.
El complejo antena funciona de la manera siguiente: cuando una de sus molculas se excita al captar un fotn (unidad
de energa lumnica) transere esa energa de excitacin a otra
molcula cercana por medio de un proceso de resonancia y,
en una reaccin en cadena, esa energa llega hasta el centro
reactivo.
Ncleo
El ncleo es un organelo caracterstico de las clulas eucariotas (g. 1-26). El material gentico de la clula se encuentra
dentro del ncleo en forma de cromatina.
El ncleo dirige las actividades de la clula, y en l tienen
lugar procesos tan importantes como la automultiplicacin del
DNA antes de comenzar la divisin celular, y la transcripcin
o produccin de los distintos tipos de RNA, que servirn para
la sntesis de protenas.
Esta estructura cambia de aspecto durante el ciclo celular
y llega a desaparecer como tal. Por ello se describe el ncleo
en interfase durante el cual es posible apreciar las siguientes
partes:
Fotosistemas
Los fotosistemas son las unidades de la membrana tilacoidal.
Cada fotosistema est formado por dos partes:
un complejo antena que consta de varios centenares de
molculas de clorola y carotenos
la envoltura nuclear, la cual est formada por dos membranas concntricas perforadas con poros nucleares; a
travs de stos se transportan molculas entre el ncleo
y el citoplasma
nucleoplasma, que es el medio interno del ncleo donde
se encuentra el resto de los componentes nucleares
nucleolo o nucleolos, los cuales son masas densas y esfricas, que constan de dos zonas: una brilar y otra granular.
ferredoxina
ESTROMA
fotn de luz
plastoquinona
fotosistema II
FAD
fotosistema I
plastocianina
ESPACIO TILACOIDAL
Figura 1-27.
ATP sintasa
Captulo 1 La clula: evolucin y desarrollo de los sistemas celulares
NADP
H+
fotn
fotn
27
H+
ATP
ADP+P
Pq
I
membrana
tilacoidal
ATP asa
II
cit b f
H+
2 H2O
O2 + 4 H
Fotolisis del agua
H+
H+
Transporte de electrones
Figura 1-28.
La brilar es interna y contiene DNA; la granular rodea
a la anterior y contiene RNA y protenas
la cromatina, constituida por DNA y protenas, aparece
durante la interfase, pero cuando la clula entra en divisin se organiza en estructuras individuales que son los
cromosomas.
Cromatina y cromosomas
Un cromosoma es una molcula de DNA muy grande que
contiene una serie de genes (g. 1-29). Un cromosoma
metafsico est formado por dos cromtides idnticas en
sentido longitudinal. En cada una de ellas hay un nucleolamento de DNA replegado idntico. Las cromtides estn
unidas por medio del centrmero. En las cromtides se aprecia
tambin un cinetocoro, centro organizador de microtbulos,
que se forma durante la mitosis y que ayuda a unir los cromosomas con el huso mittico. Por lo tanto, se puede decir
que cromatina y cromosomas es lo mismo, y el cromosoma
sera un paquete de cromatina muy compacto. Como puede
verse en la gura 1-30, en una secuencia que va desde el DNA
hasta el cromosoma.
El nmero 1 de la gura 1-30 corresponde a la molcula
de DNA; el nmero 2 ilustra el DNA unido a protenas
globulares, lo cual genera una estructura denominada collar
de perlas. Dicha estructura se origina por la repeticin de
unas unidades que son los nucleosomas. Cada nucleosoma
correspondera a una perla del collar. En el nmero 3 se pasa
a una estructura de orden superior en forma de solenoide. Se
consigue aumentar el empaquetamiento, y se genera la bra de
cromatina y nuevos bucles en el nmero 4. Hay un grado de
mayor compactacin en el nmero 5, y se establece un denso
paquete de cromatina, que es, en realidad, un cromosoma.
El total de la informacin gentica contenida en los
cromosomas de un organismo constituye su genoma.
El gen desnudo
Gen 1
Gen 2
Cromosoma
Figura 1-29. Genes.
DNA
Los genes desnudos o virus tpicos tienen 5 000 pares de
bases, es decir, 1 333 tripletes o codones y menos de 10 genes,
de manera similar a otros genes desnudos. Eso constituye
su mensaje gentico. El de la mitocondria humana tiene 38
genes. El de la Escherichia coli tiene 4 000 genes. El del Homo
sapiens tiene algo ms de 30 000 genes. A partir de estas cifras,
queda claro que los virus, viroides, fagos y plsmidos son expresiones de seres vivos muy simples. Salvo la cpside proteica
que protege, en ocasiones, a algunas de estas formas, lo ms
frecuente es encontrarlas como DNA o RNA desnudos en el
interior de una clula viva, a la cual han infectado.
28
Captulo 1
La clula: evolucin y desarrollo de los sistemas celulares
El DNA se empaca
estrechamente en los
cromosomas en metafase
5 Cromosomas
en metafase
4
3 Cromatina
condensada
Nucleosomas
1 Doble hlice
de DNA
rRNA y tRNA. Para allegarse las pocas protenas que necesitan, usan la maquinaria de rRNA y tRNA de las clulas en
cuyo interior se alojan. Los virus, viroides, etc. son piratas
(tcnicamente, parsitos) que utilizan en todo lo posible las
facilidades ya provistas en las mismas clulas vivas. Como
esa adaptacin ocurri tiempo despus de la aparicin de las
clulas vivas, se dice que las formas ms modernas de los genes
desnudos son las formas ms evolucionadas del parasitismo:
slo necesitan traer un mnimo de instrucciones para sobrevivir. Son pasajeros sin equipaje.
Al repasar el ciclo vital de un virus, se observa que, en una
etapa, la cpside llena, que ha identicado a la clula en la que
se puede hospedar, inyecta su DNA o su RNA (segn el tipo
de virus) dentro de la clula. Una de las posibilidades para el
virus de DNA es introducirse en el genoma del hospedero,
que entonces pasa de 4 000 genes a 4 010 genes. A esto se le
llama etapa lisognica.
Otra de las posibilidades para el virus de RNA o DNA es
la de utilizar la maquinaria del hospedero para multiplicarse.
Como es tan pequeo, en seguida se producen muchsimas
rplicas o copias del gen desnudo. Cada copia construye luego
su cpside proteica, y cuando los duplicados llegan a ser una
cantidad muy elevada, se produce la lisis o rotura de las membranas de la clula. Esto recibe el nombre de etapa ltica (de
rotura). De esta manera las copias quedan libres, y cada una
de ellas representa un foco de infeccin en el ambiente.
Una posibilidad ms es que un virus en etapa lisognica
sea promovido a la etapa ltica. El factor desencadenante
para ese pasaje suele ser la presencia de rayos ultravioleta,
que provienen del Sol.
Figura 1-30.
Antes de infectar a una bacteria o a cualquier otro tipo
de clula, dichos genes se visten con una cpside proteica para
dejarse llevar por el ambiente, mientras buscan su blanco a
donde llegar. La diana o blanco es la membrana de una bacteria o de una clula a la cual identican como un sitio apto
para vivir en su interior. En la gura 1-31 se observan varios
tipos de virus con su mensaje gentico protegido mediante
una cpside.
Por ser simplemente molculas de DNA o RNA, parecera que provienen de la poca de la vida del RNA, en los
albores de la existencia de la vida en el planeta.
Los virus que hoy se conocen estn adaptados por
completo (coevolucionaron) a clulas muy posteriores a la
era del RNA. Esos genes desnudos carecen de instrucciones
para fabricar ribosomas y molculas sabias, respectivamente
Tipos de virus
Adenovirus
Virus de
inmunodeficiencia
humana
Figura 1-31. Tipos de virus.
bacterifago
Captulo 1 La clula: evolucin y desarrollo de los sistemas celulares
Los virus de RNA, como es el caso del retrovirus del
SIDA, fabrica una enzima denominada retrotranscriptasa.
Con su ayuda, la molcula de RNA con todas las instrucciones virales, pasa a retrotranscribirse como DNA e, incluso,
a acceder a la etapa lisognica. El resultado es que muchas
enfermedades virales son muy difciles de curar, y algunas
parecen, hasta ahora, imposibles de ser curadas porque la
consecuencia no deseada de un proceso muy positivo, como
es la duplicacin del mensaje gentico, es que tambin se
copian las instrucciones del virus. El genoma infectado del
hospedero pasa a sus hijos y nietos, hasta que se desencadena
en un momento dado una etapa ltica, en la cual se propaga
la enfermedad a muchas clulas sanas.
Por eso se puede sospechar que fracciones mudas del
genoma con genes fragmentados son, de hecho, aunque slo
en parte, virus en etapa lisognica, que se eliminan al madurar
los genes fragmentados para dar lugar a los genes enteros y
maduros que procesan los ribosomas y las biomolculas. Si as
fuera, la maquinaria de maduracin de genes fragmentados
no sera ancestral, sino una defensa ante el reciente avance
de los genes virales, desnudos y lisognicos. Si fuese cierta
esta hiptesis, el rbol genealgico con los cinco reinos sera
menos complejo.
Un virus, entonces, es un agente gentico que posee
un cido nucleico que puede ser DNA o RNA, y que est
rodeado por una envoltura de protena. Los virus contienen
toda la informacin necesaria para su ciclo reproductor, pero
para conseguirlo necesitan a otras clulas vivas de las que
aprovechan sus organelos y molculas.
Reproduccin de los virus
La nica funcin que poseen los virus y que comparten con el
resto de los seres vivos es la de reproducirse o generar copias
de s mismos, para lo cual requieren la materia, la energa y
la maquinaria de la clula en donde se alojan, por lo que se
les denomina parsitos obligados. No poseen metabolismo ni
organizacin celular, por lo que se les sita en el lmite entre
lo vivo y lo inerte.
Una vez que los virus infectan a una clula son capaces
de manifestar dos tipos de comportamiento, bien como
agentes infecciosos que causan la lisis o muerte de la clula,
o bien como virus atenuados, que aaden material gentico
a la clula que los aloja y, por lo tanto, resultan factores de la
variabilidad gentica. Ambos casos se estudiaron con detalle
en los virus bacterifagos, y aqu se ilustra mediante unos
esquemas (g. 1-32).
Virus
Envoltura viral
Acido nucleico viral
(RNA)
Endocitosis
Endosoma
Traduccin
Ribosomas
Replicacin
RNA
viral
Transcripcin
Ncleo
RNA
Membrana
celular
Protenas
virales
Empaquetamiento
Retculo
endoplasmtico
Maduracin
Brote
Centro de ribo
nucleoprotena
Liberacin
Aparato
de Golgi
Virus
Figura 1-32. Replicacin del virus A de la influenza.
29
30
Captulo 1
La clula: evolucin y desarrollo de los sistemas celulares
En los dos casos de infeccin el proceso empieza de esta
forma:
a)
b)
c)
fase de jacin: los virus se unen por medio de la placa
basal a la cubierta de la pared bacteriana.
fase de contraccin: la cola se contrae y el cido nucleico
del virus empieza a ser inyectado.
fase de penetracin: el cido nucleico del virus penetra en
el citoplasma de la bacteria, y a partir de este momento
puede seguir dos ciclos diferentes:
en el ciclo ltico, el DNA bacteriano fabrica las protenas vricas y las copias de cidos nucleicos vricos.
Cuando hay suciente cantidad de estas molculas
se ensamblan la protena y el cido nucleico vrico,
perforan la pared celular y salen al medio, produciendo la muerte de la clula.
en el ciclo lisognico, el genoma del virus se aade
al genoma de la bacteria, no expresa sus genes y se
duplica junto al de la bacteria. El virus queda en
forma de profago.
Cuando todava los virus no revelan toda su informacin,
aparecen en el mundo microscpico otras formas acelulares de
menor tamao, capaces de causar enfermedades. Estas formas
son los viroides y los priones.
Viroides
Son molculas de RNA circular que carecen de cualquier
proteccin.
Confrmero normal
Priones
Son agentes patgenos formados por una protena (protena
del prin o PrP) que causan la enfermedad de las vacas locas o encefalopata bovina espongiforme, entre otras. Esta
protena se acumula en el cerebro de animales enfermos, lo
cual origina la estructura esponjosa de la corteza cerebral que
da nombre a la enfermedad.
Los priones, o las enfermedades producidas por priones, tienen un comportamiento sorprendente. Por un lado,
se transmiten verticalmente, como cualquier enfermedad
hereditaria caracterstica, en tanto que, por otra parte, se
comportan de manera infectiva y se transmiten de modo
horizontal mediante contagios entre individuos de distintas
especies.
La protena del prion (PrP) normal tiene una secuencia
de aminocidos (estructura primaria) idntica a la protena
del prion patgena. La diferencia entre las dos recae en las
estructuras secundaria y terciaria (g. 1-33).
La protena normal es muy rica en hlices alfa, y la
protena patgena lo es en lminas beta. Este cambio de
conformacin es crucial, ya que las protenas con lminas
beta son muy resistentes a las enzimas proteolticas, al calor
y son insolubles en agua. Pero sobre todo, la protena alterada
tiene una caracterstica nica: interacciona con una molcula
de protena normal, le cambia la conformacin y la hace capaz
de modicar las estructuras de ms protenas normales. Al
parecer, ah radica el poder infectivo de los priones.
El esquema que se presenta es muy simplicado, pues
slo se intenta explicar este proceso.
Confrmero
maligno (supuesto)
Figura 1-33. (Adaptado de http//www.cmpharmucsf.edu/cohen/research/gallery/aw_prion.gif)
Captulo 1 La clula: evolucin y desarrollo de los sistemas celulares
Puede ocurrir que la protena patgena infecte individuos
que producen protena normal (A), como ha ocurrido, por
ejemplo, cuando las vacas consumen alimento elaborado a
partir de ovejas enfermas. En este caso, la protena patgena
origina un cambio conformacional de la protena normal (B), y
transforma las hlices alfa de su estructura proteica en lminas
beta. Las nuevas protenas patgenas inducen el cambio en
otras normales, lo cual produce un efecto de cascada.
Bibliografa
Alberts B, et al. Essential cell biology. An introduction to the
molecular biology of the cell. New York/London: Garlan
Publishing, 1997.
CairnsSmith AG. Genetic takeover and the mineral origins of
life. USA: Cambridge University Press, 1982.
31
de-Duve C. Blueprint for a cell. Burlington, North Carolina: Neil
Patterson, 1991.
Ferris JP, Ertem G. Science, 1992;257:1387.
Karp G. Biologa celular y molecular. Mxico: McGrawHillInteramericana, 1998.
Lewin RA. Prochlorophyta as a proposed new division of algae.
Nature, 1976;261:697698.
Montero F, Sanz JC, Andrade MA. Evolucin prebitica: el camino
hacia la vida. Espaa: Eudema, 1993.
Reyes JJ. Vida, orden y caos. Propuestas para nuevos enfoques en biologa. Serie Libros de Texto. Mxico: Universidad Autnoma
MetropolitanaXochimilco, 1993.
Selosse MA. Les cyanobactries, dtonants procariotes photo-synthtiques. BiologieGologie, 1996;3:481.
Smith CA, Wood EJ. Molculas biolgicas. Mxico: Addison
Wesley Iberoamericana, 1997.
Solomon EP, et al. Biologa de Villee. 4a ed, Mxico: McGraw
Hill-Interamericana, 1998.
Stryer L. Bioqumica. 4a ed, Espaa: Revert, 1996. (2 tomos)
32
Captulo 1
La clula: evolucin y desarrollo de los sistemas celulares
You might also like
- Grupos FuncionalesDocument5 pagesGrupos FuncionalesLaura Sepulveda100% (1)
- Guía resueltos física mecánicaDocument7 pagesGuía resueltos física mecánicaJose Alberto100% (2)
- Estructura y función de las membranas celularesDocument6 pagesEstructura y función de las membranas celularesJuli FernandezNo ratings yet
- Comparativa FotosintesisDocument8 pagesComparativa Fotosintesisvicbaez0% (1)
- Manual de Geologia EstratosDocument19 pagesManual de Geologia Estratospagman_2006No ratings yet
- Tarea2 U4Document1 pageTarea2 U4Anonymous M5KvpjCuNo ratings yet
- Eje Funciones Logaritmicasy ExponencialesDocument1 pageEje Funciones Logaritmicasy ExponencialesAnonymous M5KvpjCuNo ratings yet
- Eje Funciones Polinomialesy RacionalesDocument2 pagesEje Funciones Polinomialesy RacionalesAnonymous M5KvpjCuNo ratings yet
- Equipo 4Document10 pagesEquipo 4Alberto JimZebNo ratings yet
- Equipo 10Document16 pagesEquipo 10Anonymous M5KvpjCuNo ratings yet
- Eje Funciones TrigonometricasDocument4 pagesEje Funciones TrigonometricasAnonymous M5KvpjCuNo ratings yet
- Eje FuncionesDocument4 pagesEje FuncionesAnonymous M5KvpjCuNo ratings yet
- Equipo 3Document6 pagesEquipo 3Anonymous M5KvpjCuNo ratings yet
- Compuestos orgánicos impacto económico industrial social ambiental ChiapasDocument8 pagesCompuestos orgánicos impacto económico industrial social ambiental ChiapasAnonymous M5KvpjCuNo ratings yet
- Equipo 4Document10 pagesEquipo 4Alberto JimZebNo ratings yet
- DiapositivasDocument9 pagesDiapositivasAnonymous M5KvpjCuNo ratings yet
- Compuestos orgánicos impacto económico industrial social ambiental ChiapasDocument8 pagesCompuestos orgánicos impacto económico industrial social ambiental ChiapasAnonymous M5KvpjCuNo ratings yet
- Equipo 1Document14 pagesEquipo 1Anonymous M5KvpjCuNo ratings yet
- Equipo 9Document12 pagesEquipo 9Anonymous M5KvpjCuNo ratings yet
- FisicaDocument21 pagesFisicaAnonymous M5KvpjCuNo ratings yet
- Equilibrio de Cuerpos (Opta)Document77 pagesEquilibrio de Cuerpos (Opta)Ivan GuerreroNo ratings yet
- Equipo 2Document7 pagesEquipo 2Anonymous M5KvpjCuNo ratings yet
- Equipo 8Document11 pagesEquipo 8Anonymous M5KvpjCuNo ratings yet
- Ejercicios Factores Conversion-Hoja 3Document2 pagesEjercicios Factores Conversion-Hoja 3Ximo Fuster100% (2)
- Equipo 7Document11 pagesEquipo 7Anonymous M5KvpjCuNo ratings yet
- Ácidos Carboxílicos del Instituto Tecnológico de Tuxtla GutiérrezDocument15 pagesÁcidos Carboxílicos del Instituto Tecnológico de Tuxtla GutiérrezAnonymous M5KvpjCuNo ratings yet
- Problemas Resueltos Cap 7 Fisica SerwayDocument14 pagesProblemas Resueltos Cap 7 Fisica Serwaylizdie71% (7)
- ESTÁTICADocument20 pagesESTÁTICASheryn Reyes PomachaguaNo ratings yet
- FisicaDocument21 pagesFisicaAnonymous M5KvpjCuNo ratings yet
- Torque, Centro de Masa y Momento AngularDocument89 pagesTorque, Centro de Masa y Momento AngularPatricia_Betan_7492100% (1)
- Cifras SignificativasDocument4 pagesCifras SignificativasAlberto JimZebNo ratings yet
- Ce09s06dc 090409054731 Phpapp02Document15 pagesCe09s06dc 090409054731 Phpapp02Anonymous M5KvpjCuNo ratings yet
- Trabajoenerga2daclase 141005094251 Conversion Gate01Document25 pagesTrabajoenerga2daclase 141005094251 Conversion Gate01Anonymous M5KvpjCuNo ratings yet
- Informe - Marcha - Analítica Grupos FuncionalesDocument5 pagesInforme - Marcha - Analítica Grupos FuncionalesMateo GavilanesNo ratings yet
- Sistemas ColoidalesDocument66 pagesSistemas ColoidalesIvanERamirezNo ratings yet
- Presentación LGNDocument13 pagesPresentación LGNYonathan SilvaNo ratings yet
- Nomenclatura orgánica fundamentalDocument16 pagesNomenclatura orgánica fundamentalOmael KingNo ratings yet
- Guia de EjerciciosDocument39 pagesGuia de EjerciciosFelicitasMilagrosNo ratings yet
- Ciclo de Vida de Alcohol A Partir de CañaDocument12 pagesCiclo de Vida de Alcohol A Partir de CañayolisjulyNo ratings yet
- Determinación de Compuestos OrgánicosDocument4 pagesDeterminación de Compuestos OrgánicosKevin Guao BolañoNo ratings yet
- Coenzima ADocument4 pagesCoenzima Aluis angello leyva riveraNo ratings yet
- HTGHRTDocument7 pagesHTGHRTMario T.CNo ratings yet
- Taller de La AtpDocument7 pagesTaller de La Atpjfvd16No ratings yet
- 5treybal 7treybal 25oconDocument7 pages5treybal 7treybal 25oconLuisa TorresNo ratings yet
- Acetona y ButanonaDocument6 pagesAcetona y ButanonaLixmelys VillarroelNo ratings yet
- Plan alimentación tilapia fertilización concentradosDocument18 pagesPlan alimentación tilapia fertilización concentradosali viloriaNo ratings yet
- Medios DiferencialesDocument1 pageMedios DiferencialesJonathan A. Carreño LaguadoNo ratings yet
- Codificacion Del ADNDocument15 pagesCodificacion Del ADNMilkobich 05No ratings yet
- Importancia del suelo y sus componentesDocument3 pagesImportancia del suelo y sus componentesAlby RamírezNo ratings yet
- Procesos 22Document39 pagesProcesos 22EDGAR ANTONIO FLORES OROSCONo ratings yet
- Práctica de Laboratorio 5Document3 pagesPráctica de Laboratorio 5David Alfaro Yufra0% (1)
- Gasolina ecológica Etanol reducir contaminación 25Document1 pageGasolina ecológica Etanol reducir contaminación 25Mila MilasNo ratings yet
- Moléculas orgánicas: carbohidratos, lípidos y proteínasDocument25 pagesMoléculas orgánicas: carbohidratos, lípidos y proteínasJuan Francisco Suárez SuárezNo ratings yet
- Bromatologia Arcos 1Document76 pagesBromatologia Arcos 1Juan Carlos Contreras Flores50% (2)
- Post BananoDocument20 pagesPost BananoRoger ContrerasNo ratings yet
- Establecimiento de Los Ejes CorporalesDocument18 pagesEstablecimiento de Los Ejes CorporalesLeonardo Emanuel Aguirre AlegriaNo ratings yet
- Los Ácidos Inorgánicos: CaracterísticasDocument7 pagesLos Ácidos Inorgánicos: CaracterísticasMaria CáceresNo ratings yet
- Es Una Molécula Compuesta Por Carbono Que Puede Adoptar Una Forma Geométrica Que Recuerda A Una EsferaDocument3 pagesEs Una Molécula Compuesta Por Carbono Que Puede Adoptar Una Forma Geométrica Que Recuerda A Una EsferaRaul HernandezNo ratings yet
- Discusión - Practica 1 - Equipo 4Document15 pagesDiscusión - Practica 1 - Equipo 4Rubi MarissaNo ratings yet