You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Biological Membranes A Biology Questions AQA OCR Edexcel PDFDocument3 pagesBiological Membranes A Biology Questions AQA OCR Edexcel PDFbekoNo ratings yet
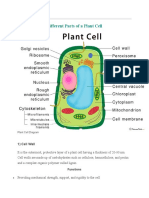
- Cell PlantDocument7 pagesCell PlantEbiz MarkNo ratings yet
- Pex 02 01Document5 pagesPex 02 01auliaNo ratings yet
- Section: Mental & Reasoning Ability: Question ID: 1034353300 Status: Not Attempted and Chosen OptionDocument46 pagesSection: Mental & Reasoning Ability: Question ID: 1034353300 Status: Not Attempted and Chosen OptionPiyush TannuNo ratings yet
- Facial ScullDocument18 pagesFacial ScullAnimesh DasNo ratings yet
- EritropoesisDocument15 pagesEritropoesisFitriyani Dewi SuwandhiNo ratings yet
- Grade 6 ScienceDocument10 pagesGrade 6 ScienceSanjeev NNo ratings yet
- Blood TransfusionDocument10 pagesBlood TransfusionDivya Joy100% (1)
- HEMA Lec 03 Bone Marrow Collection Lymphoid Organs 1Document9 pagesHEMA Lec 03 Bone Marrow Collection Lymphoid Organs 1Patrick AbellaNo ratings yet
- Biology - Zoology Unit 7 Detailed Study of FrogDocument0 pagesBiology - Zoology Unit 7 Detailed Study of Frogwww.bhawesh.com.npNo ratings yet
- Unit 7 - Respiratory SystemDocument51 pagesUnit 7 - Respiratory SystemAbdullah MohammedNo ratings yet
- FlashcardsDocument18 pagesFlashcardsDaphne HernaezNo ratings yet
- NeckDocument4 pagesNeckAttin GamerNo ratings yet
- ArchegoniateplantDocument21 pagesArchegoniateplantShubhamMalikNo ratings yet
- Laboratory Evaluation of HemostasisDocument7 pagesLaboratory Evaluation of HemostasisGerly MaglangitNo ratings yet
- Class 5 Science Classwork 091020Document3 pagesClass 5 Science Classwork 091020Rakesh AgarwalNo ratings yet
- How Do I Carry Out A Resonance Test?Document1 pageHow Do I Carry Out A Resonance Test?Inayattullah KhamkerNo ratings yet
- Anatomy and Physiology of The SkinDocument30 pagesAnatomy and Physiology of The SkinNancy VargasNo ratings yet
- Matary Surgical Anatomy 2013 AllTebFamily ComDocument320 pagesMatary Surgical Anatomy 2013 AllTebFamily Comrusu_alexandra_4No ratings yet
- Respiratory SystemDocument2 pagesRespiratory Systemlim.angelyne0126No ratings yet
- Menstrual Cycle 1Document16 pagesMenstrual Cycle 1CoxnxkNo ratings yet
- Chapter 7 - Respiratory SystemDocument24 pagesChapter 7 - Respiratory Systemchloe06muller.mNo ratings yet
- Human Anatomy: LESSON 1 Lecture NotesDocument9 pagesHuman Anatomy: LESSON 1 Lecture NotesHeikki HuuskonenNo ratings yet
- AutocoidDocument7 pagesAutocoidIdrissa ContehNo ratings yet
- Urinary System (2015 - 06 - 09 22 - 21 - 55 Utc)Document44 pagesUrinary System (2015 - 06 - 09 22 - 21 - 55 Utc)DiazNo ratings yet
- Physical Exam Vital SignsDocument4 pagesPhysical Exam Vital SignsmickeyNo ratings yet
- Grade 6 DLP Q2 W1D1Document14 pagesGrade 6 DLP Q2 W1D1CRISTINE ANN ARANDIANo ratings yet
- PHEOCHROMOCYTOMADocument28 pagesPHEOCHROMOCYTOMAQueenElsaNo ratings yet
- CBC (Complete Blood Count)Document1 pageCBC (Complete Blood Count)Roshan kumar giriNo ratings yet
- Integumentary System - NUR1019 - 20sept2022Document33 pagesIntegumentary System - NUR1019 - 20sept2022琪琪No ratings yet