You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- 6th Central Pay Commission Salary CalculatorDocument15 pages6th Central Pay Commission Salary Calculatorrakhonde100% (436)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Primary FRCA Pharmacology EquationsDocument11 pagesPrimary FRCA Pharmacology EquationsMustafa SharkawyNo ratings yet
- Primary FRCA Pharmacology EquationsDocument11 pagesPrimary FRCA Pharmacology EquationsMustafa SharkawyNo ratings yet
- Anaesthetic Drugs Cheat SheetsDocument2 pagesAnaesthetic Drugs Cheat SheetsPkern100% (3)
- The Crashing Ventilated Patient Ch.3 (Jairo I. Santanilla, ACEP, 2011) PDFDocument10 pagesThe Crashing Ventilated Patient Ch.3 (Jairo I. Santanilla, ACEP, 2011) PDFPkernNo ratings yet
- The Crashing Ventilated Patient Ch.3 (Jairo I. Santanilla, ACEP, 2011) PDFDocument10 pagesThe Crashing Ventilated Patient Ch.3 (Jairo I. Santanilla, ACEP, 2011) PDFPkernNo ratings yet
- The Crashing Ventilated Patient Ch.3 (Jairo I. Santanilla, ACEP, 2011) PDFDocument10 pagesThe Crashing Ventilated Patient Ch.3 (Jairo I. Santanilla, ACEP, 2011) PDFPkernNo ratings yet
- Selecting The Right' Positive End-Expiratory Pressure Level (Luciano Gattinoni, Current Opinion, 2016)Document8 pagesSelecting The Right' Positive End-Expiratory Pressure Level (Luciano Gattinoni, Current Opinion, 2016)PkernNo ratings yet
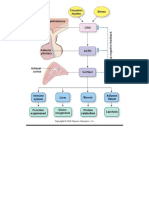
- Simple Adrenal Synthesis PathwayDocument1 pageSimple Adrenal Synthesis PathwayPkernNo ratings yet
- Respiratory-Equations (Adam Hollingworth)Document4 pagesRespiratory-Equations (Adam Hollingworth)PkernNo ratings yet
- Surgical Procedures in The Intensive Care Unit - A Critical Review. (BM Dennis, OL Gunter OA CC 2013)Document7 pagesSurgical Procedures in The Intensive Care Unit - A Critical Review. (BM Dennis, OL Gunter OA CC 2013)PkernNo ratings yet
- The Crashing Ventilated Patient Algorthm (Jairo I. Santanilla, ACEP, 2011)Document1 pageThe Crashing Ventilated Patient Algorthm (Jairo I. Santanilla, ACEP, 2011)PkernNo ratings yet
- Rule of 4 Illustration Brainstem (LITFL)Document1 pageRule of 4 Illustration Brainstem (LITFL)PkernNo ratings yet
- Spinning Dials - How To Dominate The Ventilator - Handout (EMCRIT, ARDSnet) PDFDocument5 pagesSpinning Dials - How To Dominate The Ventilator - Handout (EMCRIT, ARDSnet) PDFPkernNo ratings yet
- Human Metabolism MapDocument1 pageHuman Metabolism MapPkern83% (6)
- Heart Failure PDFDocument9 pagesHeart Failure PDFPkernNo ratings yet
- Heart FailureDocument9 pagesHeart FailurePkernNo ratings yet
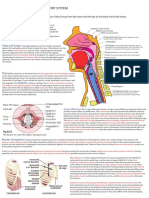
- Respiratory Notes (Chris Andersen, ICUPrimaryPrep - Com)Document14 pagesRespiratory Notes (Chris Andersen, ICUPrimaryPrep - Com)PkernNo ratings yet
- Diuretics and Renal HormonesDocument3 pagesDiuretics and Renal HormonesPkernNo ratings yet
- Pharmacodynamics and Receptor TheoryDocument8 pagesPharmacodynamics and Receptor TheoryPkernNo ratings yet
- Emergency Department Drug Card - For ALIEM - Sep 2013Document1 pageEmergency Department Drug Card - For ALIEM - Sep 2013PkernNo ratings yet
- Cardiac Murmurs and Maneuvers GuideDocument1 pageCardiac Murmurs and Maneuvers GuidePkernNo ratings yet
- STRUCTURE AND FUNCTION OF THE HEARTDocument16 pagesSTRUCTURE AND FUNCTION OF THE HEARTPkernNo ratings yet
- ED Drug Treatment Cheat Sheet ANZICSDocument1 pageED Drug Treatment Cheat Sheet ANZICSPkernNo ratings yet
- Renal, Fluids and Acid-Base Notes (Chris Andersen, ICUPrimaryPrep - Com)Document8 pagesRenal, Fluids and Acid-Base Notes (Chris Andersen, ICUPrimaryPrep - Com)PkernNo ratings yet
- Neurology Notesc (Chris Andersen, ICUPrimaryPrep - Com)Document11 pagesNeurology Notesc (Chris Andersen, ICUPrimaryPrep - Com)Pkern100% (1)
- Neurology Notesc (Chris Andersen, ICUPrimaryPrep - Com)Document11 pagesNeurology Notesc (Chris Andersen, ICUPrimaryPrep - Com)Pkern100% (1)
- 2014 CCM Review Notes Jon-Emile S. Kenny M.D, 2014Document142 pages2014 CCM Review Notes Jon-Emile S. Kenny M.D, 2014PkernNo ratings yet
- Chapter 3 The Lymphatic VasculatureDocument8 pagesChapter 3 The Lymphatic VasculaturePkernNo ratings yet
- Chapter 4 Pathophysiology of Edema FormationDocument8 pagesChapter 4 Pathophysiology of Edema FormationPkernNo ratings yet