You might also like
- Tipos de InmunidadDocument3 pagesTipos de InmunidadMari GzlezNo ratings yet
- Inmunidad CelularDocument12 pagesInmunidad CelularyaelynNo ratings yet
- InmunidadDocument7 pagesInmunidadMelanie Monserrat Sosa ArteagaNo ratings yet
- Clase Nº 06 - Del 24 Al 28 de Abril - Sistema InmunológicoDocument12 pagesClase Nº 06 - Del 24 Al 28 de Abril - Sistema InmunológicoValentina Di NoceraNo ratings yet
- Taller de InmunologiaDocument19 pagesTaller de InmunologiaGaBriiela ViDalNo ratings yet
- La Respuesta InmunitariaDocument7 pagesLa Respuesta InmunitariaJOSE ADAN NIÑO RIVASNo ratings yet
- Sistema Inmune 2020 MitreDocument9 pagesSistema Inmune 2020 Mitrelperrotta.primariaNo ratings yet
- Clase 1Document54 pagesClase 1Ana JimenezNo ratings yet
- Almagabriela - Arriaga - Unidad 1 - Actividad 2 - FORO Inmumidad InnataDocument5 pagesAlmagabriela - Arriaga - Unidad 1 - Actividad 2 - FORO Inmumidad Innatadaniel chaparroNo ratings yet
- Caracteristicas Generales Del Sistema InmunologicoDocument5 pagesCaracteristicas Generales Del Sistema InmunologicoValeria Carranza HerreraNo ratings yet
- Primera Clase InmunologiaDocument47 pagesPrimera Clase InmunologiaIvonne Mercedes Gutierrez AtapaucarNo ratings yet
- Alteraciones Del Sistema InmunológicoDocument26 pagesAlteraciones Del Sistema InmunológicoCleimar Morillo MorgadoNo ratings yet
- Inmunología Arreglada de NuevoDocument8 pagesInmunología Arreglada de NuevoJennifer De PeñaNo ratings yet
- Tarea 1 Semana 2-GUIADocument4 pagesTarea 1 Semana 2-GUIAJosefina JiménezNo ratings yet
- Respuesta InmunitariaDocument7 pagesRespuesta InmunitariaAstrid RodríguezNo ratings yet
- Monografia - Mecanismos de InmunidadDocument18 pagesMonografia - Mecanismos de InmunidadSEBASTIAN ELI VALENTIN SUNCIONNo ratings yet
- Inmunologia PreguntasDocument6 pagesInmunologia PreguntasIVONNE GERALDINE SANTACRUZ AMADORNo ratings yet
- Bases Inmunologicas de Las Vacunas-2012wDocument17 pagesBases Inmunologicas de Las Vacunas-2012wPily Williams MassNo ratings yet
- Informe 1 PDFDocument14 pagesInforme 1 PDFCristobal Severo Huamani CarrionNo ratings yet
- La InmunidadDocument3 pagesLa InmunidadMari HONo ratings yet
- Formato Word - OriginalDocument8 pagesFormato Word - Originalcarmita geronimo garciaNo ratings yet
- Respuesta Inmmune MicrobiologiaDocument7 pagesRespuesta Inmmune MicrobiologiaAndrea Gissel Mart�nez BocanegraNo ratings yet
- Farmacologia Del Sistema InmunitarioDocument23 pagesFarmacologia Del Sistema Inmunitario徳利雅83% (6)
- Cap 01Document13 pagesCap 01Fernando UsedaNo ratings yet
- Respuesta Contra MicrorganismosDocument10 pagesRespuesta Contra MicrorganismosAngeloNo ratings yet
- 02 Propiedades de La Respuesta Inmunitaria LecturaDocument14 pages02 Propiedades de La Respuesta Inmunitaria LecturamariaNo ratings yet
- Francisco Sinisterra (Codigo 2018261029) PDFDocument6 pagesFrancisco Sinisterra (Codigo 2018261029) PDFFrancisco SinisterraNo ratings yet
- InmunologíaDocument58 pagesInmunologíayeniNo ratings yet
- Sistema InmuneDocument39 pagesSistema InmuneAngelo Anaya CaponeNo ratings yet
- INMUNOLOGIADocument29 pagesINMUNOLOGIACarla LazoNo ratings yet
- Resumen de Los Capitulos de Inmunología de Faimboin 1 13 y 21Document133 pagesResumen de Los Capitulos de Inmunología de Faimboin 1 13 y 21Manuel SantellánNo ratings yet
- Control Microbiano: Respuesta InmuneDocument14 pagesControl Microbiano: Respuesta InmuneDANJHELY ALESSANDRA SANCHEZ CONDORINo ratings yet
- Cap 16 InmunologiaDocument5 pagesCap 16 InmunologiamartinvillarrealuasmedicinaNo ratings yet
- Inmunidad InnataDocument12 pagesInmunidad InnataMayra DallmannNo ratings yet
- Cuestionario InmuneDocument7 pagesCuestionario InmuneAngelica Merino100% (1)
- Respuestas InnatasDocument7 pagesRespuestas InnatasJoaquin PalaciosNo ratings yet
- Sistema InmuneDocument11 pagesSistema InmuneNicolás Polito Muñoz50% (2)
- Módulo Biología 4°medioDocument11 pagesMódulo Biología 4°medioLiceo Andrés Bello-ComunicacionesNo ratings yet
- Inmunidad InnataDocument7 pagesInmunidad Innatafatima.ponce4596No ratings yet
- 3) InmunologiaDocument29 pages3) InmunologiaJose Guillermo Mejia SaldanaNo ratings yet
- Resumen Sistema InmuneDocument6 pagesResumen Sistema Inmunekatherin_ng100% (2)
- InmunologiaDocument45 pagesInmunologiaElizabeth ArévaloNo ratings yet
- Vacunas 7 MosDocument3 pagesVacunas 7 MosBárbara OyarzoNo ratings yet
- InmunologíaDocument6 pagesInmunologíaLaura MenaNo ratings yet
- Inmunología ApuntesDocument12 pagesInmunología ApuntesclauNo ratings yet
- Causas de Las Enfermedades Por HipersensibilidadDocument8 pagesCausas de Las Enfermedades Por HipersensibilidadMayra Alejandra Navarrete AnasNo ratings yet
- Unidad 11-Inmunidad y Organos LinfaticosDocument11 pagesUnidad 11-Inmunidad y Organos LinfaticosYaz OchoaNo ratings yet
- Generalidades InmunoDocument4 pagesGeneralidades InmunoDani LópezNo ratings yet
- Libro InmunologiaDocument114 pagesLibro InmunologiaMaría Teresa Maniscalchi B.No ratings yet
- Inmuno ProyectoDocument27 pagesInmuno ProyectoMichelle YañezNo ratings yet
- Inmunidad en El Periodo NeonatalDocument30 pagesInmunidad en El Periodo NeonatalMariely GuadarramaNo ratings yet
- INMUNOLOGIADocument4 pagesINMUNOLOGIAIsabella RiveraNo ratings yet
- INMUNOLOGIADocument55 pagesINMUNOLOGIAdnfb9hsmb4No ratings yet
- Respuesta Inmune Primaria y SecundariaDocument4 pagesRespuesta Inmune Primaria y SecundariaMichelle GarciaNo ratings yet
- La Respuesta Inmunitaria RESUMENDocument4 pagesLa Respuesta Inmunitaria RESUMENDANIEL CRUZ OROZCONo ratings yet
- GL OsarioDocument1 pageGL OsarioRolando Hernandez SalazarNo ratings yet
- Ficha de Viuda NegraDocument1 pageFicha de Viuda NegraRolando Hernandez SalazarNo ratings yet
- Proteinas y TalidomidaDocument4 pagesProteinas y TalidomidaRolando Hernandez SalazarNo ratings yet
- Manual Analisis Quimicos IDocument35 pagesManual Analisis Quimicos IRolando Hernandez SalazarNo ratings yet
- Factores de Conversion de UnidadesDocument2 pagesFactores de Conversion de UnidadesDavid R PaucaraNo ratings yet
- Manual Bioquimica I Quimica Lab 2014Document51 pagesManual Bioquimica I Quimica Lab 2014Rolando Hernandez SalazarNo ratings yet
- Informe 3 AnaliticaDocument2 pagesInforme 3 AnaliticaRolando Hernandez SalazarNo ratings yet
- Apuntes de InmunoDocument103 pagesApuntes de InmunoRolando Hernandez SalazarNo ratings yet
- Matematicas 1 Trabajo FinalDocument9 pagesMatematicas 1 Trabajo FinalRolando Hernandez SalazarNo ratings yet
- Trastorno de La Personalidad AntisocialDocument1 pageTrastorno de La Personalidad AntisocialRolando Hernandez SalazarNo ratings yet
- Psicosis InfantilesDocument55 pagesPsicosis InfantilesRolando Hernandez SalazarNo ratings yet
- Instalación Del Windows Server 2003 en Una MaquinaDocument15 pagesInstalación Del Windows Server 2003 en Una MaquinaRolando Hernandez SalazarNo ratings yet
- Quimica Organica 1Document4 pagesQuimica Organica 1Rolando Hernandez SalazarNo ratings yet
- Oclusionintestinal 140329173503 Phpapp01Document92 pagesOclusionintestinal 140329173503 Phpapp01Francisco C CortesNo ratings yet
- Resumen Valoración de S DigestivoDocument18 pagesResumen Valoración de S Digestivopaula dNo ratings yet
- Aparato FonadorDocument5 pagesAparato FonadorDiliannys S.100% (1)
- Circulación AnimalDocument2 pagesCirculación AnimalMartaSoledadRamosGuillenNo ratings yet
- Reporte de Lectura #10 Cuestionario BANFE-2Document6 pagesReporte de Lectura #10 Cuestionario BANFE-2Nahid' BailyNo ratings yet
- Sistema UrinarioDocument34 pagesSistema UrinarioTalita IstefaneNo ratings yet
- Triptico RespiraciónDocument2 pagesTriptico Respiraciónjhomar45598100% (1)
- Efectos Positivos de La Actividad Física en El OrganismoDocument7 pagesEfectos Positivos de La Actividad Física en El OrganismoKevin HurtadoNo ratings yet
- Callatanzo CardioDocument210 pagesCallatanzo CardioJoaquín Aedo PozoNo ratings yet
- Endocrinología NemotecniasDocument3 pagesEndocrinología Nemotecniasalbertohz10% (1)
- Mapa Mental SDocument1 pageMapa Mental SMitzi HerreraNo ratings yet
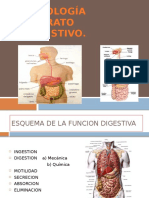
- Fisiologia Del Aparato DigestivoDocument34 pagesFisiologia Del Aparato DigestivoIlianPlacencia100% (1)
- Puntos Dolorosos AbdominalesDocument2 pagesPuntos Dolorosos AbdominalesJhonny SalinasNo ratings yet
- Sistema LinfáticoDocument7 pagesSistema LinfáticoAnthon Esquivel100% (1)
- (William H Fitzgerald) Reflexología Podálica - Masaje Zonal en Los PiesDocument24 pages(William H Fitzgerald) Reflexología Podálica - Masaje Zonal en Los PiesBelkis Albertt86% (7)
- AP. CirculatorioDocument15 pagesAP. CirculatorioKatiAriasNo ratings yet
- Salinotas Embrio 1P - 230126 - 135442 - 230629 - 165550Document28 pagesSalinotas Embrio 1P - 230126 - 135442 - 230629 - 165550Melissa RangelNo ratings yet
- Topografía General Del Peritoneo Del AdultoDocument6 pagesTopografía General Del Peritoneo Del AdultoMercedes RoblesNo ratings yet
- Mano Acupuntura CoreanaDocument488 pagesMano Acupuntura Coreanamvil2397% (95)
- Cap. 5Document16 pagesCap. 5Sequeira Jz JohnNo ratings yet
- Mapa Conceptual Del Sistema EndrocrinoDocument1 pageMapa Conceptual Del Sistema EndrocrinoLorena Rivero89% (9)
- Desarrollo Embrionario Y Diferenciacion Sexual: 1.1. SexoDocument19 pagesDesarrollo Embrionario Y Diferenciacion Sexual: 1.1. SexojuanNo ratings yet
- Traumatismo TesticularDocument10 pagesTraumatismo Testicularisabela camacho garciaNo ratings yet
- Actividad Ciencias Naturales 7moDocument1 pageActividad Ciencias Naturales 7moMia AztridNo ratings yet
- Mapa Mapas MergedDocument3 pagesMapa Mapas MergedEmily MontesNo ratings yet
- 4radiologiacabezaycuello 110426193206 Phpapp02 PDFDocument52 pages4radiologiacabezaycuello 110426193206 Phpapp02 PDFEstefania MoralesNo ratings yet
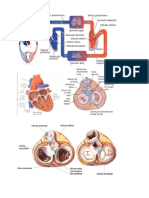
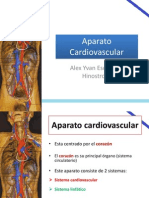
- Anatomía de Tórax: Aparato Cardiovascular (Corazón)Document78 pagesAnatomía de Tórax: Aparato Cardiovascular (Corazón)Alex Yvan Escobedo Hinostroza100% (1)
- Banco de PreguntasDocument22 pagesBanco de PreguntasAlex Aguilar GarciaNo ratings yet
- Marco de Referencia Control MotorDocument21 pagesMarco de Referencia Control MotorMaría Fernanda Villaseca0% (1)
- Folleto Sistema EndocrinoDocument5 pagesFolleto Sistema EndocrinoSindy SantamariaNo ratings yet
- Recupera tu mente, reconquista tu vidaFrom EverandRecupera tu mente, reconquista tu vidaRating: 5 out of 5 stars5/5 (6)
- El Monje Que Vendio Su Ferrari: Una Fábula EspiritualFrom EverandEl Monje Que Vendio Su Ferrari: Una Fábula EspiritualRating: 4.5 out of 5 stars4.5/5 (1698)
- Tus Zonas Erroneas: Guía Para Combatir las Causas de la InfelicidadFrom EverandTus Zonas Erroneas: Guía Para Combatir las Causas de la InfelicidadRating: 4.5 out of 5 stars4.5/5 (1831)
- Psicología oscura: Una guía esencial de persuasión, manipulación, engaño, control mental, negociación, conducta humana, PNL y guerra psicológicaFrom EverandPsicología oscura: Una guía esencial de persuasión, manipulación, engaño, control mental, negociación, conducta humana, PNL y guerra psicológicaRating: 4.5 out of 5 stars4.5/5 (766)
- Tu cerebro emocional: Saca partido de lo que sientes y transforma tu vidaFrom EverandTu cerebro emocional: Saca partido de lo que sientes y transforma tu vidaRating: 5 out of 5 stars5/5 (2)
- Cómo hacer que te pasen cosas buenas: Entiende tu cerebro, gestiona tus emociones, mejora tu vidaFrom EverandCómo hacer que te pasen cosas buenas: Entiende tu cerebro, gestiona tus emociones, mejora tu vidaRating: 5 out of 5 stars5/5 (1870)
- La revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaFrom EverandLa revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaRating: 5 out of 5 stars5/5 (201)
- Yo Pude, ¡Tú Puedes!: Cómo tomar el control de tu bienestar emocional y convertirte en una persona imparable (edición revisada y expandida)From EverandYo Pude, ¡Tú Puedes!: Cómo tomar el control de tu bienestar emocional y convertirte en una persona imparable (edición revisada y expandida)Rating: 5 out of 5 stars5/5 (7)
- Cómo Acertar en tu Nueva Relación de Pareja: Todas las claves para saber si tu relación tiene futuro... o te conviene salir corriendo de ahí.From EverandCómo Acertar en tu Nueva Relación de Pareja: Todas las claves para saber si tu relación tiene futuro... o te conviene salir corriendo de ahí.Rating: 4.5 out of 5 stars4.5/5 (7)
- Resetea tu mente. Descubre de lo que eres capazFrom EverandResetea tu mente. Descubre de lo que eres capazRating: 5 out of 5 stars5/5 (196)
- Los Secretos De La Mente Millonaria: Domina el juego de la riquezaFrom EverandLos Secretos De La Mente Millonaria: Domina el juego de la riquezaRating: 5 out of 5 stars5/5 (457)
- ¡Tómate un respiro! Mindfulness: El arte de mantener la calma en medio de la tempestadFrom Everand¡Tómate un respiro! Mindfulness: El arte de mantener la calma en medio de la tempestadRating: 5 out of 5 stars5/5 (198)
- ¿Por qué mis padres no me aman?: Empezando a sanarFrom Everand¿Por qué mis padres no me aman?: Empezando a sanarRating: 4.5 out of 5 stars4.5/5 (33)
- Resumen de Pensar rápido pensar despacio de Daniel KahnemanFrom EverandResumen de Pensar rápido pensar despacio de Daniel KahnemanRating: 4.5 out of 5 stars4.5/5 (64)
- Homo antecessor: El nacimiento de una especieFrom EverandHomo antecessor: El nacimiento de una especieRating: 5 out of 5 stars5/5 (1)
- Mindfulness para principiantes: Medita sin meditarFrom EverandMindfulness para principiantes: Medita sin meditarRating: 5 out of 5 stars5/5 (53)
- Disciplina Mental: Técnicas infalibles para lograr todo lo que te propones y eliminar la pereza y la procrastinación de tu vida para siempreFrom EverandDisciplina Mental: Técnicas infalibles para lograr todo lo que te propones y eliminar la pereza y la procrastinación de tu vida para siempreRating: 5 out of 5 stars5/5 (3)
- El poder del optimismo: Herramientas para vivir de forma más positivaFrom EverandEl poder del optimismo: Herramientas para vivir de forma más positivaRating: 5 out of 5 stars5/5 (16)
- Influencia. La psicología de la persuasiónFrom EverandInfluencia. La psicología de la persuasiónRating: 4.5 out of 5 stars4.5/5 (14)
- Grimorio: Cómo lanzar y elaborar hechizos mágicos, aprender las prácticas wiccanas y desvelar los secretos de la brujería a través de un diario ritualFrom EverandGrimorio: Cómo lanzar y elaborar hechizos mágicos, aprender las prácticas wiccanas y desvelar los secretos de la brujería a través de un diario ritualNo ratings yet
- Cómo terminar lo que empiezas: El arte de perseverar, pasar a la acción, ejecutar los planes y tener disciplinaFrom EverandCómo terminar lo que empiezas: El arte de perseverar, pasar a la acción, ejecutar los planes y tener disciplinaRating: 4.5 out of 5 stars4.5/5 (6)
- Teoría polivagal práctica y terapiaFrom EverandTeoría polivagal práctica y terapiaRating: 5 out of 5 stars5/5 (4)