You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- WtWf-Trin AberrantTeragenDocument144 pagesWtWf-Trin AberrantTeragenRichard DeRemer50% (2)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Cro-Magnon Man-Part 8-RaemonDocument19 pagesCro-Magnon Man-Part 8-Raemonapi-242202307No ratings yet
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Ethiopian HistoryDocument144 pagesEthiopian HistoryDagninet Yamlak100% (1)
- Social Anthropology FinalDocument62 pagesSocial Anthropology FinalKumera Dinkisa Tolera100% (1)
- Human Evolution and The Origins of Hierarchies The State of NatureDocument288 pagesHuman Evolution and The Origins of Hierarchies The State of NatureGeorge Alex100% (1)
- Evolution The Human StoryDocument258 pagesEvolution The Human StoryEjder Yapar96% (47)
- Act 1.2 Evolution of ManDocument3 pagesAct 1.2 Evolution of ManRay FaustinoNo ratings yet
- Tabon Cave excavations uncover early human fossils in the PhilippinesDocument4 pagesTabon Cave excavations uncover early human fossils in the PhilippinesJan Arlyn Juanico100% (1)
- Ina Wunn - Beginning of ReligionDocument36 pagesIna Wunn - Beginning of ReligionRob ThuhuNo ratings yet
- Merlin Donald, Origins of The Modern Mind PDFDocument425 pagesMerlin Donald, Origins of The Modern Mind PDFjarubirubi92% (12)
- Close Encounters of The Prehistoric Kind: OnlineDocument5 pagesClose Encounters of The Prehistoric Kind: Onlinejohnny cartinNo ratings yet
- Homo Erectus4Document18 pagesHomo Erectus4johnny cartinNo ratings yet
- 723 (2010) Hernán A. Burbano,: Science Et AlDocument4 pages723 (2010) Hernán A. Burbano,: Science Et Aljohnny cartinNo ratings yet
- A Draft Sequence of The Neandertal Genome (Sapiens/Neanderthal Interbreeding)Document14 pagesA Draft Sequence of The Neandertal Genome (Sapiens/Neanderthal Interbreeding)José Pedro GomesNo ratings yet
- Homo Erectus2Document2 pagesHomo Erectus2johnny cartinNo ratings yet
- Cell, Vol. 90, 19-30, July 11, 1997, Copyright ©1997 byDocument12 pagesCell, Vol. 90, 19-30, July 11, 1997, Copyright ©1997 byjohnny cartinNo ratings yet
- Gorillas in CrisisDocument2 pagesGorillas in Crisisjohnny cartinNo ratings yet
- A Cranium For The Earliest Europeans: Phylogenetic Position of The Hominid From Ceprano, ItalyDocument6 pagesA Cranium For The Earliest Europeans: Phylogenetic Position of The Hominid From Ceprano, Italyjohnny cartinNo ratings yet
- Ts17 Gorilla EDocument84 pagesTs17 Gorilla Ejohnny cartinNo ratings yet
- Earliest Evidence of Modern Human Life History in North AfricanDocument6 pagesEarliest Evidence of Modern Human Life History in North Africanjohnny cartinNo ratings yet
- Neanader 2 PDFDocument5 pagesNeanader 2 PDFjohnny cartinNo ratings yet
- Homo Erectus1Document4 pagesHomo Erectus1johnny cartinNo ratings yet
- Inconsistencies in Neanderthal Genomic DNA SequencesDocument5 pagesInconsistencies in Neanderthal Genomic DNA Sequencesjohnny cartinNo ratings yet
- Neanader 2 PDFDocument5 pagesNeanader 2 PDFjohnny cartinNo ratings yet
- Ieidelbergensis 1Document10 pagesIeidelbergensis 1johnny cartinNo ratings yet
- A Cranium For The Earliest Europeans: Phylogenetic Position of The Hominid From Ceprano, ItalyDocument6 pagesA Cranium For The Earliest Europeans: Phylogenetic Position of The Hominid From Ceprano, Italyjohnny cartinNo ratings yet
- Erg Aster 1Document12 pagesErg Aster 1johnny cartinNo ratings yet
- A New Skull of Early Homo From Dmanisi, Georgia AbesalomDocument6 pagesA New Skull of Early Homo From Dmanisi, Georgia Abesalomjohnny cartinNo ratings yet
- Erg Aster 4Document8 pagesErg Aster 4johnny cartinNo ratings yet
- Foraging of Homo Ergaster and Australopithecus Boisei in East African EnvironmentsDocument6 pagesForaging of Homo Ergaster and Australopithecus Boisei in East African Environmentsjohnny cartinNo ratings yet
- The First Female Homo Erectus Pelvis, From Gona, Afar, EthiopiaDocument5 pagesThe First Female Homo Erectus Pelvis, From Gona, Afar, Ethiopiajohnny cartinNo ratings yet
- Annali Dell'Università Degli Studi Di Ferrara Museologia Scientifica e NaturalisticaDocument5 pagesAnnali Dell'Università Degli Studi Di Ferrara Museologia Scientifica e Naturalisticajohnny cartinNo ratings yet
- Controversial Human Ancestor Gets Major Facelift - LiveScienceDocument3 pagesControversial Human Ancestor Gets Major Facelift - LiveSciencejohnny cartinNo ratings yet
- Ann. Zoot. Fennici 28:37/-380 Helsi Nki 19 February 1992Document10 pagesAnn. Zoot. Fennici 28:37/-380 Helsi Nki 19 February 1992johnny cartinNo ratings yet
- American Journal of Physical Anthropology 138:90-100 (2009)Document11 pagesAmerican Journal of Physical Anthropology 138:90-100 (2009)johnny cartinNo ratings yet
- American Journal of Physical Anthropology 138:90-100 (2009)Document11 pagesAmerican Journal of Physical Anthropology 138:90-100 (2009)johnny cartinNo ratings yet
- Chapter B The Great Ice AgeDocument2 pagesChapter B The Great Ice Agejohnny cartinNo ratings yet
- Ouranopithecus MacedoniensisDocument27 pagesOuranopithecus Macedoniensisjohnny cartinNo ratings yet
- Journal of Human Evolution 46 (2004) 433-465Document33 pagesJournal of Human Evolution 46 (2004) 433-465johnny cartinNo ratings yet
- Human Evolution: Full ImageDocument2 pagesHuman Evolution: Full Imagenikita bajpaiNo ratings yet
- 1999-Tattersall - Neandertalci U Ljudskoj EvolucijiDocument3 pages1999-Tattersall - Neandertalci U Ljudskoj EvolucijiPavla MackicNo ratings yet
- 2019 Haplogroup RDocument713 pages2019 Haplogroup RKlaus MarklNo ratings yet
- DioramaDocument9 pagesDioramaChris Kirk Patrick PatweckNo ratings yet
- Mitochondrial DNA Rare in E Europe and N Asia PDFDocument12 pagesMitochondrial DNA Rare in E Europe and N Asia PDFAlexander HagenNo ratings yet
- Soal Tes Kemampuan Dan Potensi Akademik SNPTN2019Document7 pagesSoal Tes Kemampuan Dan Potensi Akademik SNPTN2019Yollyver KotarumalosNo ratings yet
- 2.2 - Tabon CavesDocument1 page2.2 - Tabon CavesTrishNo ratings yet
- Unit One Introducing Anthropology and Its Subject Matter 1.1 Definition, Scope and Subject Matter of AnthropologyDocument14 pagesUnit One Introducing Anthropology and Its Subject Matter 1.1 Definition, Scope and Subject Matter of Anthropologyfeyeko aberaNo ratings yet
- EvolutionDocument16 pagesEvolutionAagash PranavNo ratings yet
- Human Evolution Timeline: Homo Habilis Homo Neanderthalensis: AkaDocument1 pageHuman Evolution Timeline: Homo Habilis Homo Neanderthalensis: Akacykabylat3087No ratings yet
- Anthropology SCERT KERALA CLASS-11@Pdf4examsDocument343 pagesAnthropology SCERT KERALA CLASS-11@Pdf4examsShiv Shakti SinghNo ratings yet
- Research PaperDocument5 pagesResearch Paperapi-241126921No ratings yet
- NOVA Evolution Labs Missions 4-6Document15 pagesNOVA Evolution Labs Missions 4-6DENZEL ORTIZNo ratings yet
- Hublin Et Al 2017 African Origin H Sapiens MoroccoDocument17 pagesHublin Et Al 2017 African Origin H Sapiens MoroccoRodrigo Giesta FigueiredoNo ratings yet
- ANTHROPOLOGY 101: WHAT IS ANTHROPOLOGYDocument42 pagesANTHROPOLOGY 101: WHAT IS ANTHROPOLOGYNatnael MisrakNo ratings yet
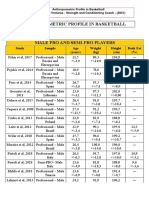
- Basketball: Anthropometric Profile of PlayersDocument6 pagesBasketball: Anthropometric Profile of PlayersAdriano VretarosNo ratings yet
- Evolution and Progress of Early ManDocument5 pagesEvolution and Progress of Early ManZiaUlhaqNo ratings yet
- Human 20evolutionDocument17 pagesHuman 20evolutionArchit GulkariNo ratings yet
- Boas, General AnthropologyDocument730 pagesBoas, General AnthropologyPepe TruenoNo ratings yet
- Syl Woo AnthDocument9 pagesSyl Woo AnthWajahat GhafoorNo ratings yet