You might also like
- Consciousness: Theories in Neuroscience and Philosophy of MindFrom EverandConsciousness: Theories in Neuroscience and Philosophy of MindNo ratings yet
- Intuition Part1Document12 pagesIntuition Part1Luis Alberto Rebaza DominguezNo ratings yet
- Gale Researcher Guide for: Experimental and Behavioral PsychologyFrom EverandGale Researcher Guide for: Experimental and Behavioral PsychologyNo ratings yet
- HeartMath Inuition StudyDocument12 pagesHeartMath Inuition StudyAdrian AlecuNo ratings yet
- Connecting Consciousness: The Role of Large Scale Brain Networks in ConsciousnessDocument33 pagesConnecting Consciousness: The Role of Large Scale Brain Networks in ConsciousnessHariNo ratings yet
- Massimini PBR Coma Science 2009Document14 pagesMassimini PBR Coma Science 2009hamletquironNo ratings yet
- Gillett 1989Document21 pagesGillett 1989sadek boughezalaNo ratings yet
- Erro Ahmad Surya 1611900027 Academic Writing ADocument5 pagesErro Ahmad Surya 1611900027 Academic Writing AErro SuryaNo ratings yet
- The Brain ConsciousnessDocument25 pagesThe Brain ConsciousnessImtiaz Bashir100% (1)
- Consciousness Origins of PDFDocument6 pagesConsciousness Origins of PDF11111No ratings yet
- Sam Parnia - Reports of Consciousness During Cardiac Arrest PDFDocument5 pagesSam Parnia - Reports of Consciousness During Cardiac Arrest PDFChris SchelinNo ratings yet
- Mind The Brain Mind The Brain Mind The Brain Mind The BrainDocument4 pagesMind The Brain Mind The Brain Mind The Brain Mind The BrainMariø YŋgveøhliŋNo ratings yet
- CONSCIOUSNESS IN THE UNIVERSE AN UPDATED REVIEW OF THE "ORCH OR" THEORY Stuart R. Hameroff, and Roger PenroseDocument114 pagesCONSCIOUSNESS IN THE UNIVERSE AN UPDATED REVIEW OF THE "ORCH OR" THEORY Stuart R. Hameroff, and Roger Penrosecrisolaris100% (1)
- Emotion and The Human Brain: Antonio R. DamasioDocument6 pagesEmotion and The Human Brain: Antonio R. DamasioBruno XavierNo ratings yet
- Manifesto For A Post Materialist ScienceDocument4 pagesManifesto For A Post Materialist ScienceMichel BlancNo ratings yet
- Neuroscience and Saint Thomas AquinasDocument3 pagesNeuroscience and Saint Thomas AquinasAnonymous Vld2tfTAJCNo ratings yet
- Raymond Trevor Bradley - The Psychophysiology of Intuition: A Quantum-Holographic Theory of Nonlocal CommunicationDocument38 pagesRaymond Trevor Bradley - The Psychophysiology of Intuition: A Quantum-Holographic Theory of Nonlocal CommunicationDominos021100% (1)
- Relația Dintre Creier Și Conștiință: (Popper, 1978 Crick, 1994 Libet, 2004) (Libet, 2004) (Singer, 2015)Document3 pagesRelația Dintre Creier Și Conștiință: (Popper, 1978 Crick, 1994 Libet, 2004) (Libet, 2004) (Singer, 2015)Mihaela ElaNo ratings yet
- 4E CognitionDocument8 pages4E CognitionCoteOlivaPuginNo ratings yet
- Robert G. Jahn and Brenda J. Dunne - A Modular Model of Mind/Matter Manifestations (M 5)Document31 pagesRobert G. Jahn and Brenda J. Dunne - A Modular Model of Mind/Matter Manifestations (M 5)Dominos021No ratings yet
- The Nature of PrecognitionDocument20 pagesThe Nature of Precognitionmauricio gómez100% (1)
- Editorial: J.Neurol Sci (Turk)Document4 pagesEditorial: J.Neurol Sci (Turk)Nezih OktarNo ratings yet
- Psychology Chapters Review1Document14 pagesPsychology Chapters Review1Tabita PetreanNo ratings yet
- Brain ActivityDocument11 pagesBrain Activityanon_457484602No ratings yet
- Consciousness Quantum TheoryDocument12 pagesConsciousness Quantum TheoryMokhtar MohdNo ratings yet
- Conscious ConnectionDocument17 pagesConscious ConnectionProf VRShanbhagNo ratings yet
- Mechanisms For TopDown Perceptual ProcessingDocument15 pagesMechanisms For TopDown Perceptual ProcessingGeetika RaoNo ratings yet
- The Brain Is Not Mental! Coupling Neuronal and Immune Cellular Processing in Human OrganismsDocument13 pagesThe Brain Is Not Mental! Coupling Neuronal and Immune Cellular Processing in Human OrganismsLiaNo ratings yet
- The Entropic Brain A Theory of Conscious States Informed by Neuroimaging Research With Psychedelic DrugsDocument22 pagesThe Entropic Brain A Theory of Conscious States Informed by Neuroimaging Research With Psychedelic DrugsFabricio García FragosoNo ratings yet
- Notes - Psychokinesis - From Jack Houck's Website. Pioneered PK PartiesDocument10 pagesNotes - Psychokinesis - From Jack Houck's Website. Pioneered PK PartiesVas RaNo ratings yet
- Biology of Consciousness PDFDocument7 pagesBiology of Consciousness PDFMano RatyiNo ratings yet
- Incidence of Neuroscience On The Psychopedagogy Relation Corporeity/learningDocument11 pagesIncidence of Neuroscience On The Psychopedagogy Relation Corporeity/learningjona_josephNo ratings yet
- 3 Seth Consciousness ResearchDocument2 pages3 Seth Consciousness ResearchNatalia MenottiNo ratings yet
- MerkerDocument60 pagesMerkerscribd_new_accountNo ratings yet
- A Study On Consumer Perception Towards Life Insurance in Satna CityDocument64 pagesA Study On Consumer Perception Towards Life Insurance in Satna CityAbhay Jain100% (1)
- Emotion Review 2013 Panksepp 235 40Document7 pagesEmotion Review 2013 Panksepp 235 40Anca PrajanuNo ratings yet
- The Claustrum and Consciousness - An UpdateDocument12 pagesThe Claustrum and Consciousness - An UpdateAnton PatriaNo ratings yet
- Le VanQuyen - Neurodynamics and Phenomenology in Mutual Enlightenment: The Example of The Epileptic Aura.Document25 pagesLe VanQuyen - Neurodynamics and Phenomenology in Mutual Enlightenment: The Example of The Epileptic Aura.goo99No ratings yet
- White Paper XXVDocument21 pagesWhite Paper XXVCefasSimonNo ratings yet
- Anderson 2006 Cognitive Science and Epistemic OpennessDocument41 pagesAnderson 2006 Cognitive Science and Epistemic Opennesssmac226No ratings yet
- Consumer Perception Towards LIC in Rewa CityDocument64 pagesConsumer Perception Towards LIC in Rewa CityAbhay JainNo ratings yet
- 0262011816.MIT Press - Sunny Y. Auyang - Life, Mind in Everyday & Science, Cognitive - May.2009Document526 pages0262011816.MIT Press - Sunny Y. Auyang - Life, Mind in Everyday & Science, Cognitive - May.2009Branko NikolicNo ratings yet
- 10.1017 s0140525x00012759 PDFDocument62 pages10.1017 s0140525x00012759 PDFkristian FelipeNo ratings yet
- Mashour 2013Document8 pagesMashour 2013Carolina LuditzaNo ratings yet
- Interoception and Emotion - A Neuroanatomical PerspectiveDocument22 pagesInteroception and Emotion - A Neuroanatomical PerspectiveYuki MatsutaniNo ratings yet
- Biology of Consciousness: Gerald M. Edelman, Joseph A. Gally and Bernard J. BaarsDocument7 pagesBiology of Consciousness: Gerald M. Edelman, Joseph A. Gally and Bernard J. Baarskfun90No ratings yet
- Somatosensory Processes Subserving Perception and ActionDocument51 pagesSomatosensory Processes Subserving Perception and ActionGustavo CabanasNo ratings yet
- Quantum Mechanics of 'Conscious Energy'Document29 pagesQuantum Mechanics of 'Conscious Energy'SYED ISMYL MAHMOOD RIZVINo ratings yet
- Experiential Fantasies Prediction and en PDFDocument26 pagesExperiential Fantasies Prediction and en PDFIvan MatancevicNo ratings yet
- Emotion Systems Cells Synaptic PlasticitDocument7 pagesEmotion Systems Cells Synaptic PlasticitLeonardo PunshonNo ratings yet
- Schwartz Consciousness Spirituality and Postmaterialist ScienceDocument14 pagesSchwartz Consciousness Spirituality and Postmaterialist ScienceSeanNo ratings yet
- ContentServer PDFDocument16 pagesContentServer PDFredcloud111No ratings yet
- Consciousness As A Concrete Physical PhenomenonDocument12 pagesConsciousness As A Concrete Physical PhenomenonthalesmmsNo ratings yet
- Event Horizons of The PsycheDocument234 pagesEvent Horizons of The PsycheDavid Bruce Albert Jr., Ph.D.100% (2)
- Nonlinear Dynamics of The Brain: Emotion and Cognition: Home Search Collections Journals About Contact Us My IopscienceDocument17 pagesNonlinear Dynamics of The Brain: Emotion and Cognition: Home Search Collections Journals About Contact Us My Iopsciencebrinkman94No ratings yet
- States of Consciousness 3Document4 pagesStates of Consciousness 3renate_ellaNo ratings yet
- Articulo Lisa FeldmanDocument28 pagesArticulo Lisa FeldmanLaia Colom BertrånNo ratings yet
- Miracles in The BrainDocument10 pagesMiracles in The BrainIVAN ALVAREZNo ratings yet
- Dick Bierman - Do PSI-phenomena Suggest Radical Dualism?Document7 pagesDick Bierman - Do PSI-phenomena Suggest Radical Dualism?Canola_OliveNo ratings yet
- M. Hohol Urbanczyk Embodied Embedded SocCog LibreDocument20 pagesM. Hohol Urbanczyk Embodied Embedded SocCog LibreCopernicus Center for Interdisciplinary StudiesNo ratings yet
- 2 Bad BricksDocument3 pages2 Bad Bricksalxzndr__No ratings yet
- Feelings Are SufferingDocument1 pageFeelings Are Sufferingalxzndr__No ratings yet
- Forum: The: Lojong J JNG NGDocument12 pagesForum: The: Lojong J JNG NGalxzndr__100% (1)
- MIDI-OX 7.0: Installation NotesDocument7 pagesMIDI-OX 7.0: Installation Notesalxzndr__No ratings yet
- MIDIOX Change HistoryDocument10 pagesMIDIOX Change Historyalxzndr__No ratings yet
- Mox ScriptDocument12 pagesMox Scriptalxzndr__100% (1)
- Isabel Clarke Psychosis and SpiritualityDocument5 pagesIsabel Clarke Psychosis and Spiritualityalxzndr__No ratings yet
- Sound GuideDocument2 pagesSound Guidealxzndr__No ratings yet
- Student Management System - Full DocumentDocument46 pagesStudent Management System - Full DocumentI NoNo ratings yet
- Title: Daily Visit Report: SDL Mini - Project Academic Year 2020-21 Group ID:GB6Document2 pagesTitle: Daily Visit Report: SDL Mini - Project Academic Year 2020-21 Group ID:GB6Arjuna JppNo ratings yet
- ESS Revision Session 2 - Topics 5-8 & P1 - 2Document54 pagesESS Revision Session 2 - Topics 5-8 & P1 - 2jinLNo ratings yet
- Civil Engineering Interview QuestionsDocument19 pagesCivil Engineering Interview QuestionsSrivardhanSrbNo ratings yet
- Soul Winners' SecretsDocument98 pagesSoul Winners' Secretsmichael olajideNo ratings yet
- 03-Volume II-A The MIPS64 Instruction Set (MD00087)Document793 pages03-Volume II-A The MIPS64 Instruction Set (MD00087)miguel gonzalezNo ratings yet
- Surat Textile MillsDocument3 pagesSurat Textile MillsShyam J VyasNo ratings yet
- Harper 2001Document6 pagesHarper 2001Elena GologanNo ratings yet
- United States Court of Appeals, Third CircuitDocument8 pagesUnited States Court of Appeals, Third CircuitScribd Government DocsNo ratings yet
- Didhard Muduni Mparo and 8 Others Vs The GRN of Namibia and 6 OthersDocument20 pagesDidhard Muduni Mparo and 8 Others Vs The GRN of Namibia and 6 OthersAndré Le RouxNo ratings yet
- Dialogue About Handling ComplaintDocument3 pagesDialogue About Handling ComplaintKarimah Rameli100% (4)
- Byron and The Bulgarian Revival Period - Vitana KostadinovaDocument7 pagesByron and The Bulgarian Revival Period - Vitana KostadinovavitanaNo ratings yet
- Honey ProcessingDocument5 pagesHoney Processingvenkatrao_gvNo ratings yet
- Latvian Adjectives+Document6 pagesLatvian Adjectives+sherin PeckalNo ratings yet
- Rotation and Revolution of EarthDocument4 pagesRotation and Revolution of EarthRamu ArunachalamNo ratings yet
- Maratua Island Survey ReportDocument8 pagesMaratua Island Survey ReportJoko TrisyantoNo ratings yet
- ZultaniteDocument4 pagesZultaniteAcharya BalwantNo ratings yet
- أثر البحث والتطوير على النمو الاقتصادي - دراسة قياسية لحالة الجزائر (1990 -2014)Document17 pagesأثر البحث والتطوير على النمو الاقتصادي - دراسة قياسية لحالة الجزائر (1990 -2014)Star FleurNo ratings yet
- IsaiahDocument7 pagesIsaiahJett Rovee Navarro100% (1)
- Skylab Our First Space StationDocument184 pagesSkylab Our First Space StationBob AndrepontNo ratings yet
- Kerjaya JuruterbangDocument11 pagesKerjaya JuruterbangAqil NazriNo ratings yet
- Research On Goat Nutrition and Management in Mediterranean Middle East and Adjacent Arab Countries IDocument20 pagesResearch On Goat Nutrition and Management in Mediterranean Middle East and Adjacent Arab Countries IDebraj DattaNo ratings yet
- Hoc Volume1Document46 pagesHoc Volume1nordurljosNo ratings yet
- For Printing Week 5 PerdevDocument8 pagesFor Printing Week 5 PerdevmariNo ratings yet
- Design and Experimental Performance Assessment of An Outer Rotor PM Assisted SynRM For The Electric Bike PropulsionDocument11 pagesDesign and Experimental Performance Assessment of An Outer Rotor PM Assisted SynRM For The Electric Bike PropulsionTejas PanchalNo ratings yet
- Test Bank For Cognitive Psychology Connecting Mind Research and Everyday Experience 3rd Edition e Bruce GoldsteinDocument24 pagesTest Bank For Cognitive Psychology Connecting Mind Research and Everyday Experience 3rd Edition e Bruce GoldsteinMichaelThomasyqdi100% (49)
- 2nd Quarter Exam All Source g12Document314 pages2nd Quarter Exam All Source g12Bobo Ka100% (1)
- Law - Midterm ExamDocument2 pagesLaw - Midterm ExamJulian Mernando vlogsNo ratings yet
- Pencak Silat New Rules 2020 - Slides Presentation (International) - As of 22 Aug 2020 - 1000hrs (1) (201-400)Document200 pagesPencak Silat New Rules 2020 - Slides Presentation (International) - As of 22 Aug 2020 - 1000hrs (1) (201-400)Yasin ilmansyah hakimNo ratings yet
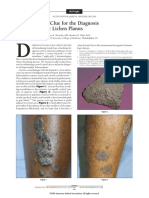
- A Novel Visual Clue For The Diagnosis of Hypertrophic Lichen PlanusDocument1 pageA Novel Visual Clue For The Diagnosis of Hypertrophic Lichen Planus600WPMPONo ratings yet
- The Age of Magical Overthinking: Notes on Modern IrrationalityFrom EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityRating: 4 out of 5 stars4/5 (30)
- ADHD is Awesome: A Guide to (Mostly) Thriving with ADHDFrom EverandADHD is Awesome: A Guide to (Mostly) Thriving with ADHDRating: 5 out of 5 stars5/5 (3)
- The Stoic Mindset: Living the Ten Principles of StoicismFrom EverandThe Stoic Mindset: Living the Ten Principles of StoicismRating: 4.5 out of 5 stars4.5/5 (11)
- The 7 Habits of Highly Effective People: The Infographics EditionFrom EverandThe 7 Habits of Highly Effective People: The Infographics EditionRating: 4 out of 5 stars4/5 (2475)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedRating: 5 out of 5 stars5/5 (81)
- Love Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)From EverandLove Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)Rating: 3 out of 5 stars3/5 (1)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeFrom EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeRating: 2 out of 5 stars2/5 (1)
- Master Your Emotions: Develop Emotional Intelligence and Discover the Essential Rules of When and How to Control Your FeelingsFrom EverandMaster Your Emotions: Develop Emotional Intelligence and Discover the Essential Rules of When and How to Control Your FeelingsRating: 4.5 out of 5 stars4.5/5 (321)
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionFrom EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionRating: 4 out of 5 stars4/5 (404)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsFrom EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsNo ratings yet
- The One Thing: The Surprisingly Simple Truth Behind Extraordinary ResultsFrom EverandThe One Thing: The Surprisingly Simple Truth Behind Extraordinary ResultsRating: 4.5 out of 5 stars4.5/5 (709)
- Indistractable: How to Control Your Attention and Choose Your LifeFrom EverandIndistractable: How to Control Your Attention and Choose Your LifeRating: 3 out of 5 stars3/5 (5)
- The Power of Now: A Guide to Spiritual EnlightenmentFrom EverandThe Power of Now: A Guide to Spiritual EnlightenmentRating: 4.5 out of 5 stars4.5/5 (4125)
- Summary: Atomic Habits by James Clear: An Easy & Proven Way to Build Good Habits & Break Bad OnesFrom EverandSummary: Atomic Habits by James Clear: An Easy & Proven Way to Build Good Habits & Break Bad OnesRating: 5 out of 5 stars5/5 (1636)
- Becoming Supernatural: How Common People Are Doing The UncommonFrom EverandBecoming Supernatural: How Common People Are Doing The UncommonRating: 5 out of 5 stars5/5 (1482)
- The 5 Second Rule: Transform your Life, Work, and Confidence with Everyday CourageFrom EverandThe 5 Second Rule: Transform your Life, Work, and Confidence with Everyday CourageRating: 5 out of 5 stars5/5 (12)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsFrom EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsRating: 5 out of 5 stars5/5 (1)
- Summary: The Laws of Human Nature: by Robert Greene: Key Takeaways, Summary & AnalysisFrom EverandSummary: The Laws of Human Nature: by Robert Greene: Key Takeaways, Summary & AnalysisRating: 4.5 out of 5 stars4.5/5 (30)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessFrom EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessRating: 4.5 out of 5 stars4.5/5 (328)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsFrom EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsRating: 4 out of 5 stars4/5 (4)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaFrom EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaRating: 4.5 out of 5 stars4.5/5 (266)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (5)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryFrom EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryRating: 4 out of 5 stars4/5 (46)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.From EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Rating: 4.5 out of 5 stars4.5/5 (110)