You might also like
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Locus Coeruleus Noradrenergic - Topics by Science - GovDocument211 pagesLocus Coeruleus Noradrenergic - Topics by Science - GovRehaan FayazNo ratings yet
- PNAS Gene Knock Out PDFDocument6 pagesPNAS Gene Knock Out PDFRehaan FayazNo ratings yet
- Aston-Jones & Cohen (ARN 05)Document50 pagesAston-Jones & Cohen (ARN 05)Rehaan FayazNo ratings yet
- aLL ABOUT cATECHOLAMINES SYNTHESIS N REUPTAKEDocument72 pagesaLL ABOUT cATECHOLAMINES SYNTHESIS N REUPTAKERehaan FayazNo ratings yet
- Issues and Opinions in Nutrition: Utility and Importance of Gene Knockout Animals For Nutritional and Metabolic ResearchDocument6 pagesIssues and Opinions in Nutrition: Utility and Importance of Gene Knockout Animals For Nutritional and Metabolic ResearchRehaan FayazNo ratings yet
- Chapter 1 - Real NumbersDocument19 pagesChapter 1 - Real NumberskdNo ratings yet
- 01 Basics of Computers PDFDocument19 pages01 Basics of Computers PDFRehaan FayazNo ratings yet
- Sustainable Development Goals (SDGS)Document4 pagesSustainable Development Goals (SDGS)Rehaan FayazNo ratings yet
- Welfare Schemes For Women in IndiaDocument2 pagesWelfare Schemes For Women in Indialoga priyaNo ratings yet
- 1st Sssion On Regulations23052018byFSSAIDocument20 pages1st Sssion On Regulations23052018byFSSAIRehaan FayazNo ratings yet
- Government Schemes in India: Scheme Name Launch Sector DetailsDocument8 pagesGovernment Schemes in India: Scheme Name Launch Sector DetailsRehaan FayazNo ratings yet
- DBTBaramulla PDFDocument24 pagesDBTBaramulla PDFRehaan FayazNo ratings yet
- FAQs ImportDocument10 pagesFAQs ImportRehaan FayazNo ratings yet
- Student World Atlas Malestrom PDFDocument115 pagesStudent World Atlas Malestrom PDFMeisterKurosawaNo ratings yet
- International Headquarters Organisation Headquarters Head/Secretary GeneralDocument7 pagesInternational Headquarters Organisation Headquarters Head/Secretary GeneralRehaan FayazNo ratings yet
- Current Affaird-April-2020Document90 pagesCurrent Affaird-April-2020Abhishek AswalNo ratings yet
- UPSC Prelims Solution 2016 by GKTodayDocument26 pagesUPSC Prelims Solution 2016 by GKTodayasef grgrNo ratings yet
- AssignmentDocument5 pagesAssignmentRehaan FayazNo ratings yet
- Sleep Deprivation Induced by The Modified Multiple Platform Technique PDFDocument7 pagesSleep Deprivation Induced by The Modified Multiple Platform Technique PDFRehaan FayazNo ratings yet
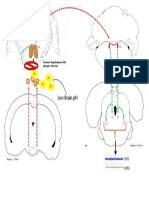
- VLPO, TMN, LC InteractionDocument1 pageVLPO, TMN, LC InteractionRehaan FayazNo ratings yet
- Women and Armed Conflict: Widows in Kashmir': International Journal of Sociology and AnthropologyDocument8 pagesWomen and Armed Conflict: Widows in Kashmir': International Journal of Sociology and AnthropologyRehaan FayazNo ratings yet
- Review Article: Extracellular Vesicles and Their Convergence With Viral PathwaysDocument13 pagesReview Article: Extracellular Vesicles and Their Convergence With Viral PathwaysRehaan FayazNo ratings yet
- UPSC IFS Botany SyllabusDocument3 pagesUPSC IFS Botany SyllabusRehaan FayazNo ratings yet
- NN 2344Document8 pagesNN 2344Rehaan FayazNo ratings yet
- J. Biol. Chem.-2004-Askwith-18296-305Document11 pagesJ. Biol. Chem.-2004-Askwith-18296-305Rehaan FayazNo ratings yet
- DNA MethylationDocument22 pagesDNA MethylationRehaan FayazNo ratings yet
- Sleep Alterations in MammalsDocument13 pagesSleep Alterations in MammalsRehaan FayazNo ratings yet
- Niti AyogDocument4 pagesNiti AyogkkrrkumarNo ratings yet
- Dreadd Tech1Document36 pagesDreadd Tech1Rehaan FayazNo ratings yet
- 100 Questions On EnvironmentDocument20 pages100 Questions On EnvironmentvgasNo ratings yet
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Molinspiration ChembioinformaticsDocument6 pagesMolinspiration ChembioinformaticsbedibediNo ratings yet
- Advances in Stem Cell ResearchDocument400 pagesAdvances in Stem Cell ResearchbluegreenalgaNo ratings yet
- 1 s2.0 S2772782322000249 MainDocument11 pages1 s2.0 S2772782322000249 MainAbdelmounaim El KhattabiNo ratings yet
- 2004 June Module 1Document11 pages2004 June Module 1navin-10No ratings yet
- TMP D820Document9 pagesTMP D820FrontiersNo ratings yet
- World of Corn 2005Document10 pagesWorld of Corn 2005National Corn Growers AssociationNo ratings yet
- PCR Lab Write UpDocument4 pagesPCR Lab Write Upmadypaddie50% (2)
- ZnT8 Nature GeneticsDocument28 pagesZnT8 Nature GeneticsDeepak JainNo ratings yet
- Recombinant Vaccine DevelopmentDocument29 pagesRecombinant Vaccine DevelopmentShivraj JadhavNo ratings yet
- Desai 2008Document3 pagesDesai 2008Raul GamiñoNo ratings yet
- Study and Work in Sweden: A GuideDocument28 pagesStudy and Work in Sweden: A GuideSoonia100% (16)
- Gmo 1Document45 pagesGmo 1Meziel LamanilaoNo ratings yet
- Essay On ChildrenDocument6 pagesEssay On Childrenb71bpjha100% (2)
- Syllabus UN3501 GU4501 20180822Document11 pagesSyllabus UN3501 GU4501 20180822Katherine XuNo ratings yet
- (Medical Radiology) Jonathan Strauss, William Small JR., Gayle E. Woloschak - Breast Cancer Biology For The Radiation Oncologist-Springer (2015)Document94 pages(Medical Radiology) Jonathan Strauss, William Small JR., Gayle E. Woloschak - Breast Cancer Biology For The Radiation Oncologist-Springer (2015)Piotr JankowskiNo ratings yet
- BIOCHEMISTRY CHAPTER 1 INTRODUCTION With CONCEPT MAPDocument4 pagesBIOCHEMISTRY CHAPTER 1 INTRODUCTION With CONCEPT MAPKASHMIR R3No ratings yet
- Galactose Metabolism in Saccharomyces CerevisiaeDocument12 pagesGalactose Metabolism in Saccharomyces CerevisiaeFarida RahayuNo ratings yet
- Concise Histology - Leslie P Gartner & James L Hiatt (1E)Document353 pagesConcise Histology - Leslie P Gartner & James L Hiatt (1E)Subhashree Nayak94% (16)
- HW - Carbohydrate Metabolism II & Lipid MetabolismDocument2 pagesHW - Carbohydrate Metabolism II & Lipid MetabolismyanNo ratings yet
- DNA Replication Concepts and ConnectionsDocument7 pagesDNA Replication Concepts and ConnectionslolasparkleNo ratings yet
- RT-PCR Test Detects No COVID-19Document1 pageRT-PCR Test Detects No COVID-19rubabshaikhNo ratings yet
- Covid-19 PCR Test Report for Mr. Sidharth SwaroopDocument1 pageCovid-19 PCR Test Report for Mr. Sidharth SwaroopSidharth SwaroopNo ratings yet
- Cellular Transport Test Review ExplainedDocument4 pagesCellular Transport Test Review ExplainedJustin Tis0% (1)
- Otagiri, 2016. Albumin in Medicine. EBOOKDocument279 pagesOtagiri, 2016. Albumin in Medicine. EBOOKFinda Istiqomah100% (1)
- General Biology Act NotebookDocument2 pagesGeneral Biology Act NotebookSheene MariquinaNo ratings yet
- Laboratory Manuel - Application of Molecular Methods in Microbiology, Biochemistry and Plant PhysiologyDocument26 pagesLaboratory Manuel - Application of Molecular Methods in Microbiology, Biochemistry and Plant PhysiologyiuventasNo ratings yet
- LIF101 Mid Semester ExaminationDocument6 pagesLIF101 Mid Semester ExaminationPiyush KumarNo ratings yet
- Microbial Biotechnology in The Laboratory and Practice Theory, ExercisesDocument553 pagesMicrobial Biotechnology in The Laboratory and Practice Theory, ExercisesRamya KrishnamurthyNo ratings yet
- The CellDocument13 pagesThe Cellsl07021No ratings yet
- MicrometabolismDocument41 pagesMicrometabolismMohammed Mansour AbdullahNo ratings yet