You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Diagnosing Anemia in Neonates An Evidence-Based ApproachDocument13 pagesDiagnosing Anemia in Neonates An Evidence-Based Approacharmando salvador100% (1)
- Johannes Gerbes, Frauke Van Der Werff - Fit Fürs Goethe-Zertifikat, A2 - Start Deutsch 2, Volume 2 (2007, Hueber Verlag)Document33 pagesJohannes Gerbes, Frauke Van Der Werff - Fit Fürs Goethe-Zertifikat, A2 - Start Deutsch 2, Volume 2 (2007, Hueber Verlag)kareemNo ratings yet
- What Is The Oxygen CascadeDocument27 pagesWhat Is The Oxygen CascadeRandhir Singh90% (10)
- British Columbia (Public Service Employee ReDocument32 pagesBritish Columbia (Public Service Employee ReChristine GildensternNo ratings yet
- Test Bank For Wilkins Clinical Assessment in Respiratory Care 8th Edition HeuerDocument15 pagesTest Bank For Wilkins Clinical Assessment in Respiratory Care 8th Edition Heueraaronraymd15012001cew100% (14)
- Bittner 2015 Acute and Perioperative Care of TheDocument34 pagesBittner 2015 Acute and Perioperative Care of TheJuan FelipeNo ratings yet
- BHS InggrisDocument9 pagesBHS InggrisAies AishaNo ratings yet
- 01 - Blood-Function and CompositionDocument45 pages01 - Blood-Function and Compositionsylvester GelacNo ratings yet
- Important Questions On Bioinorganic Chemistry 80Document7 pagesImportant Questions On Bioinorganic Chemistry 80Fahad KhanNo ratings yet
- The Bohr EffectDocument16 pagesThe Bohr EffectZaid SulimanNo ratings yet
- Interpreting Laboratory Results: Key PointsDocument8 pagesInterpreting Laboratory Results: Key PointsMarjorie Joy GreciaNo ratings yet
- Urine Physical ExamDocument3 pagesUrine Physical ExamFait HeeNo ratings yet
- Urine AnalysisDocument3 pagesUrine AnalysisFara Liyana RahimNo ratings yet
- Drugs Acting On Blood and Blood Forming AgentsDocument29 pagesDrugs Acting On Blood and Blood Forming AgentsSri RamNo ratings yet
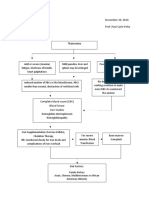
- Concept MapDocument2 pagesConcept MapcheskaotreraNo ratings yet
- BiochemistryDocument68 pagesBiochemistry20AR018 HARIHARA SUBRAMANIANNo ratings yet
- Environmental and Occupational ToxicantsDocument3 pagesEnvironmental and Occupational ToxicantsEhrli CuaresmaNo ratings yet
- Questions Bank For Molecular Biology Answer Sheet PDFDocument51 pagesQuestions Bank For Molecular Biology Answer Sheet PDFsajjad hussainNo ratings yet
- The Sequence of Events That Occur in The Heart During Cardiac CycleDocument13 pagesThe Sequence of Events That Occur in The Heart During Cardiac CycleADITYAROOP PATHAKNo ratings yet
- Physiology, Oxygen Transport: April 2019Document5 pagesPhysiology, Oxygen Transport: April 2019Soha SonaNo ratings yet
- Biology 2023 U2 P2Document19 pagesBiology 2023 U2 P2johnathanNo ratings yet
- 123Document4 pages123JC Cayanan0% (1)
- Haemoglobinopathies - ThalassaemiaDocument49 pagesHaemoglobinopathies - ThalassaemiaRaymond LundaNo ratings yet
- 1 - Toxic GasesDocument19 pages1 - Toxic GasesxIRONxWOLFxNo ratings yet
- Palm Vein TechnologyDocument26 pagesPalm Vein TechnologyRakshitha RinkuNo ratings yet
- Function of The Respiratory SystemDocument5 pagesFunction of The Respiratory SystemRachelHatcher123No ratings yet
- Sickle Cell AnemiaDocument10 pagesSickle Cell Anemiaaitzaz ul haqNo ratings yet
- Enzymology NotesDocument5 pagesEnzymology NotesRyan Fortune AludaNo ratings yet
- B PDFDocument31 pagesB PDFNazaqat FarooqNo ratings yet
- Blood MCQDocument32 pagesBlood MCQHashim Omar91% (11)