You might also like
- Multi-Nutrient Blocks en Goats Feeding, in Vitro Evaluation, Navarro Et Al. 2020Document15 pagesMulti-Nutrient Blocks en Goats Feeding, in Vitro Evaluation, Navarro Et Al. 2020Emmanuel A. Sessarego DávilaNo ratings yet
- Mammalian Sex Reversal and Intersexuality, 2000Document7 pagesMammalian Sex Reversal and Intersexuality, 2000Emmanuel A. Sessarego DávilaNo ratings yet
- Frequency of Disease-Causing Genetic Mutations in Mixed-Breed Dogs, Zierath Et Al. 2017Document13 pagesFrequency of Disease-Causing Genetic Mutations in Mixed-Breed Dogs, Zierath Et Al. 2017Emmanuel A. Sessarego DávilaNo ratings yet
- Dog Coat Colour Genetics - A Review, Saif Et Al. 2020Document11 pagesDog Coat Colour Genetics - A Review, Saif Et Al. 2020Emmanuel A. Sessarego DávilaNo ratings yet
- Immunological Properties of Donkey Milk - Its Potential Use in The Prevention of Atherosclerosis, Tafaro Et Al. 2007Document8 pagesImmunological Properties of Donkey Milk - Its Potential Use in The Prevention of Atherosclerosis, Tafaro Et Al. 2007Emmanuel A. Sessarego DávilaNo ratings yet
- Lysozyme Activity in Donkey Milk, Martini Et Al. 2019Document4 pagesLysozyme Activity in Donkey Milk, Martini Et Al. 2019Emmanuel A. Sessarego DávilaNo ratings yet
- Macroanatomical Aspects of Gastrointestinal Tract of Alpaca and Dromedary, Pérez Et Al. 2016Document7 pagesMacroanatomical Aspects of Gastrointestinal Tract of Alpaca and Dromedary, Pérez Et Al. 2016Emmanuel A. Sessarego DávilaNo ratings yet
- Animal NutritionDocument714 pagesAnimal NutritionZozo Mostafa92% (12)
- Whey Proteins and Their Antimicrobial Properties in Donkey Milk: A Brief ReviewDocument15 pagesWhey Proteins and Their Antimicrobial Properties in Donkey Milk: A Brief ReviewMaria AspriNo ratings yet
- A Web Resource On DNA Tests For Canine and Feline Hereditary Diseases, Slutsky Et Al. 2013Document6 pagesA Web Resource On DNA Tests For Canine and Feline Hereditary Diseases, Slutsky Et Al. 2013EMMANUEL ALEXANDER SESSAREGO DÁVILANo ratings yet
- Frequency of Disease-Causing Genetic Mutations in Mixed-Breed Dogs, Zierath Et Al. 2017Document13 pagesFrequency of Disease-Causing Genetic Mutations in Mixed-Breed Dogs, Zierath Et Al. 2017Emmanuel A. Sessarego DávilaNo ratings yet
- Dog Coat Colour Genetics - A Review, Saif Et Al. 2020Document11 pagesDog Coat Colour Genetics - A Review, Saif Et Al. 2020Emmanuel A. Sessarego DávilaNo ratings yet
- Lysozyme Activity in Donkey Milk, Martini Et Al. 2019Document4 pagesLysozyme Activity in Donkey Milk, Martini Et Al. 2019Emmanuel A. Sessarego DávilaNo ratings yet
- Morphological Charactirization of Amiata Donkey Through Multivariate Analyses, Sargentini Et Al. 2018Document10 pagesMorphological Charactirization of Amiata Donkey Through Multivariate Analyses, Sargentini Et Al. 2018Emmanuel A. Sessarego DávilaNo ratings yet
- Sarti2019 Article DonkeySMilkInTheManagementOfCh PDFDocument9 pagesSarti2019 Article DonkeySMilkInTheManagementOfCh PDFMaria AspriNo ratings yet
- Donkey Milk Proteins - Digestibility and Nutritional Significance, Marletta Et Al. 2016Document12 pagesDonkey Milk Proteins - Digestibility and Nutritional Significance, Marletta Et Al. 2016Emmanuel A. Sessarego DávilaNo ratings yet
- Sarti2019 Article DonkeySMilkInTheManagementOfCh PDFDocument9 pagesSarti2019 Article DonkeySMilkInTheManagementOfCh PDFMaria AspriNo ratings yet
- Morphological Charactirization of Amiata Donkey Through Multivariate Analyses, Sargentini Et Al. 2018Document10 pagesMorphological Charactirization of Amiata Donkey Through Multivariate Analyses, Sargentini Et Al. 2018Emmanuel A. Sessarego DávilaNo ratings yet
- Donkey Milk Proteins - Digestibility and Nutritional Significance, Marletta Et Al. 2016Document12 pagesDonkey Milk Proteins - Digestibility and Nutritional Significance, Marletta Et Al. 2016Emmanuel A. Sessarego DávilaNo ratings yet
- Strategies to Improve Donkey Semen FertilityDocument7 pagesStrategies to Improve Donkey Semen FertilityEmmanuel A. Sessarego DávilaNo ratings yet
- Tolerability of Donkey Milk in 92 Highly-Problematic Cow Milk Allergic Children, Monti Et Al. 2012Document8 pagesTolerability of Donkey Milk in 92 Highly-Problematic Cow Milk Allergic Children, Monti Et Al. 2012Emmanuel A. Sessarego DávilaNo ratings yet
- The Antimicrobial Activity of Donkey Milk and Its Microflora Changes During Storage, Xiao-Ying Et Al. 2008Document5 pagesThe Antimicrobial Activity of Donkey Milk and Its Microflora Changes During Storage, Xiao-Ying Et Al. 2008Emmanuel A. Sessarego DávilaNo ratings yet
- The Antimicrobial Activity of Donkey Milk and Its Microflora Changes During Storage, Xiao-Ying Et Al. 2008Document5 pagesThe Antimicrobial Activity of Donkey Milk and Its Microflora Changes During Storage, Xiao-Ying Et Al. 2008Emmanuel A. Sessarego DávilaNo ratings yet
- Lapte Magarita 3Document9 pagesLapte Magarita 3Ontica CameliaNo ratings yet
- Lapte Magarita 3Document9 pagesLapte Magarita 3Ontica CameliaNo ratings yet
- Antimicrobial Effect of Donkey Milk Digested in Vitro Qith Human Gastrointestinal Enzymes, Tidona Et Al. 2011 PDFDocument8 pagesAntimicrobial Effect of Donkey Milk Digested in Vitro Qith Human Gastrointestinal Enzymes, Tidona Et Al. 2011 PDFEmmanuel A. Sessarego DávilaNo ratings yet
- Whey Proteins and Their Antimicrobial Properties in Donkey Milk: A Brief ReviewDocument15 pagesWhey Proteins and Their Antimicrobial Properties in Donkey Milk: A Brief ReviewMaria AspriNo ratings yet
- Tolerability of Donkey Milk in 92 Highly-Problematic Cow Milk Allergic Children, Monti Et Al. 2012Document8 pagesTolerability of Donkey Milk in 92 Highly-Problematic Cow Milk Allergic Children, Monti Et Al. 2012Emmanuel A. Sessarego DávilaNo ratings yet
- Strategies to Improve Donkey Semen FertilityDocument7 pagesStrategies to Improve Donkey Semen FertilityEmmanuel A. Sessarego DávilaNo ratings yet
- Whey Proteins and Their Antimicrobial Properties in Donkey Milk: A Brief ReviewDocument15 pagesWhey Proteins and Their Antimicrobial Properties in Donkey Milk: A Brief ReviewMaria AspriNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Poultry Production Egg Weight (G) : 1 Animal ScienceDocument109 pagesPoultry Production Egg Weight (G) : 1 Animal ScienceAlvin WatinNo ratings yet
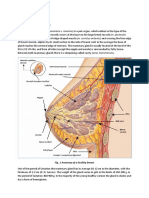
- Breast anatomy overviewDocument59 pagesBreast anatomy overviewgina2535100% (1)
- Impaired Lactation Review of Delayed Lactogenesis and Insufficient LactationDocument10 pagesImpaired Lactation Review of Delayed Lactogenesis and Insufficient LactationRob BordenNo ratings yet
- SonoSim Objectives BreastDocument8 pagesSonoSim Objectives Breastkingjameson1No ratings yet
- Breastfeeding: BY: Dinabandhu Barad MSC Tutor Dept. of Child Health Nursing SNC, Soa, DtuDocument57 pagesBreastfeeding: BY: Dinabandhu Barad MSC Tutor Dept. of Child Health Nursing SNC, Soa, DtuAbdul Azis G. CamidNo ratings yet
- LACTATION Physiology and ManagementDocument37 pagesLACTATION Physiology and ManagementRed Williams0% (2)
- Apoyo A Diada - Guia para SLPDocument11 pagesApoyo A Diada - Guia para SLPPaula Belén Rojas CunazzaNo ratings yet
- Understanding the Normal Puerperium ProcessDocument52 pagesUnderstanding the Normal Puerperium ProcessPiyush Dutta100% (1)
- Dairy Chemistry and Biochemistry by P. F. Fox and P. L. H. McSweeneyDocument495 pagesDairy Chemistry and Biochemistry by P. F. Fox and P. L. H. McSweeneySMITKUMAR PATEL100% (1)
- ChapterDocument5 pagesChapterAyu WiliariNo ratings yet
- The Unitarian or Trophoblastic Thesis of CancerDocument32 pagesThe Unitarian or Trophoblastic Thesis of Canceruge9No ratings yet
- Lesson 05. Mammary GlandsDocument23 pagesLesson 05. Mammary GlandsLola PNo ratings yet
- Mammary GlandsDocument6 pagesMammary GlandsRendy SusantoNo ratings yet
- Biology MCQs and Short QuestionsDocument20 pagesBiology MCQs and Short QuestionsKeshav MehtaNo ratings yet
- Seminar On Physiology of PuerperiumDocument15 pagesSeminar On Physiology of Puerperiumvarshasharma05100% (3)
- Female Reproductive HistologyDocument59 pagesFemale Reproductive HistologyIta Indriani100% (2)
- ThesisDocument108 pagesThesisvasantvj100% (2)
- Anatomy and Physiology of the BreastDocument3 pagesAnatomy and Physiology of the BreastOrtezz RachNo ratings yet
- Bovine Mastitis SVDocument62 pagesBovine Mastitis SVSatyavrat SinghNo ratings yet
- Pregnancy, Lactation, InfancyDocument32 pagesPregnancy, Lactation, InfancyCheska YsabelleNo ratings yet
- Definition and Stages of LactationDocument27 pagesDefinition and Stages of LactationDakshayini MbNo ratings yet
- 4 1Document265 pages4 1sillypoloNo ratings yet
- Breast AnatomyDocument8 pagesBreast AnatomyHossamuldin Aly El NemrNo ratings yet
- BREAST Development TSADocument62 pagesBREAST Development TSAshirley_ling_15No ratings yet
- Breast Cancer Term PaperDocument86 pagesBreast Cancer Term Papersusy mary thomasNo ratings yet
- Unit 5 Normal Puerperium and Nursing Management: 5.0 ObjectivesDocument14 pagesUnit 5 Normal Puerperium and Nursing Management: 5.0 ObjectivesJissa Daison100% (1)
- Aniscan Dairy ScienceDocument23 pagesAniscan Dairy Scienceishumathi13No ratings yet
- Agriculture Form 4 NotesDocument24 pagesAgriculture Form 4 Notesbirgenenock47No ratings yet
- Cells of The Human BodyDocument633 pagesCells of The Human BodyAshley Diane Henry100% (1)
- Physiology Passmrcog 2016Document73 pagesPhysiology Passmrcog 2016Mona HelouNo ratings yet