You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Unit 5 Lab Questions ForestryDocument2 pagesUnit 5 Lab Questions ForestryClara GreenNo ratings yet
- Soal Latihan Kelas 9Document7 pagesSoal Latihan Kelas 9Ika Yuniati WinataNo ratings yet
- Deepwater Horizon Oil Spill: Science, Law & PolicyDocument28 pagesDeepwater Horizon Oil Spill: Science, Law & PolicyNational Press FoundationNo ratings yet
- 003 SD 01Document7 pages003 SD 01Patricia Nicole ElediaNo ratings yet
- Listening Sample Task 9 PDFDocument3 pagesListening Sample Task 9 PDFDana ContrasNo ratings yet
- The Third World War Will Be Fought ForDocument3 pagesThe Third World War Will Be Fought ForNeha RastogiNo ratings yet
- LESSON PLAN Water ConservationDocument8 pagesLESSON PLAN Water ConservationkarlacamilledeleonNo ratings yet
- TRF Case Study SheetDocument1 pageTRF Case Study Sheetapi-253104811No ratings yet
- MapsDocument12 pagesMapsdonna benitoNo ratings yet
- IUCN - Stung Treng Policy Brief 2 - Integrated Assessment of Wetlands in CambodiaDocument4 pagesIUCN - Stung Treng Policy Brief 2 - Integrated Assessment of Wetlands in Cambodiadavid_allen3028No ratings yet
- AssignmentDocument5 pagesAssignmentParadox SkyLarkNo ratings yet
- Comments 1417439984Document32 pagesComments 1417439984api-64666718No ratings yet
- 'Mystery Dinner Snake' Found in Belly of Another Snake Finally IdentifiedDocument2 pages'Mystery Dinner Snake' Found in Belly of Another Snake Finally IdentifiedGina PiñeroNo ratings yet
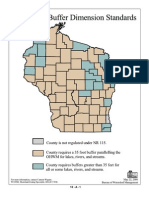
- Shoreland Vegetation - Buffer - Standards by County in WisconsinDocument29 pagesShoreland Vegetation - Buffer - Standards by County in WisconsinLori KoschnickNo ratings yet
- Mountain Pygmy-Possum Burramys Action StatementDocument5 pagesMountain Pygmy-Possum Burramys Action Statementapi-328095923No ratings yet
- Irrigation Objective Type Question and AnswersDocument13 pagesIrrigation Objective Type Question and Answersprabu061100% (3)
- Barrage PresentationDocument10 pagesBarrage Presentationpinky100% (1)
- Client StoryDocument1 pageClient StorybernabethNo ratings yet
- Black Cottonwood and Balsam Poplar Managers' Handbook For British ColumbiaDocument126 pagesBlack Cottonwood and Balsam Poplar Managers' Handbook For British ColumbiaCarlsen RockwellNo ratings yet
- Materia SecaDocument12 pagesMateria SecaCristian Alejandro Vásquez TituanaNo ratings yet
- Rehabilitation of Perungudi DumpyardDocument4 pagesRehabilitation of Perungudi DumpyardAkshayaNo ratings yet
- How To Protect Mother EarthDocument4 pagesHow To Protect Mother EarthJeevanya Janani RanjithNo ratings yet
- Mining Pollution: The Case of The Baguio Mining District, The PhilippinesDocument10 pagesMining Pollution: The Case of The Baguio Mining District, The PhilippinesFritz GuevarraNo ratings yet
- Environment An Outline of The Causes of Deforestation in Cambodia PDFDocument7 pagesEnvironment An Outline of The Causes of Deforestation in Cambodia PDFmilky calditNo ratings yet
- Wildlife Conservation Efforts in IndiaDocument2 pagesWildlife Conservation Efforts in Indiashresthaprashant8006No ratings yet
- Worksheets ITnCT EngDocument21 pagesWorksheets ITnCT EngneocorpuzNo ratings yet
- Environment Law Project FinalDocument23 pagesEnvironment Law Project FinalAmarendraKumar100% (1)
- Dhyum The DugongDocument32 pagesDhyum The DugongStrawbearyNo ratings yet
- BeanBiodiversity Jatun Sacha ReadyDocument7 pagesBeanBiodiversity Jatun Sacha ReadyJohn OsborneNo ratings yet
- Chapter 5 - Geologic AgentsDocument28 pagesChapter 5 - Geologic AgentsfiqNo ratings yet