You might also like
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- CEO Administrator Healthcare Management in San Antonio TX Resume Robert WardDocument3 pagesCEO Administrator Healthcare Management in San Antonio TX Resume Robert WardRobertWard2No ratings yet
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Art of Healthy Eating KidsDocument122 pagesThe Art of Healthy Eating KidsSenka SkenderovicNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Herbal Medicine: Current Status and The FutureDocument8 pagesHerbal Medicine: Current Status and The FuturexiuhtlaltzinNo ratings yet
- Clinical Case in Contact Lenses, 1e - 2001Document205 pagesClinical Case in Contact Lenses, 1e - 2001Miguel PalaciosNo ratings yet
- Drug Study DengueDocument3 pagesDrug Study DengueiamELHIZANo ratings yet
- BLS Adult Skills Checklist 2016Document2 pagesBLS Adult Skills Checklist 2016Wahyudi QorahmanNo ratings yet
- DLP in MAPEH - Grade 9 Myths and Misconceptions About DrugsDocument4 pagesDLP in MAPEH - Grade 9 Myths and Misconceptions About DrugsMa. Reina Gail T. Lizaso100% (5)
- Fanconi AnemiaDocument2 pagesFanconi AnemiaAreef MuarifNo ratings yet
- Progeria SyndromeDocument1 pageProgeria SyndromeAreef MuarifNo ratings yet
- Sotos Syndrome / Cerebral Gigantism /: Clinical SynopsisDocument2 pagesSotos Syndrome / Cerebral Gigantism /: Clinical SynopsisAreef MuarifNo ratings yet
- Alagille SyndromeDocument2 pagesAlagille SyndromeAreef MuarifNo ratings yet
- VATER AssociationDocument2 pagesVATER AssociationAreef MuarifNo ratings yet
- Rett Syndrome RTT /: OMIM2008 Clinical Synopsis Clinical Synopsis Clinical Synopsis Clinical SynopsisDocument2 pagesRett Syndrome RTT /: OMIM2008 Clinical Synopsis Clinical Synopsis Clinical Synopsis Clinical SynopsisAreef MuarifNo ratings yet
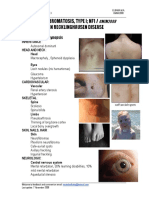
- NeurofibromatosisType IDocument2 pagesNeurofibromatosisType IAreef MuarifNo ratings yet
- Cri Du Chat SyndromeDocument2 pagesCri Du Chat SyndromeAreef MuarifNo ratings yet
- Noonan SyndromeDocument3 pagesNoonan SyndromeAreef MuarifNo ratings yet
- Silver Rassell SyndromeDocument2 pagesSilver Rassell SyndromeAreef MuarifNo ratings yet
- Auditory Processing and Cognitive Profiles in Children with Specific Learning DisorderDocument9 pagesAuditory Processing and Cognitive Profiles in Children with Specific Learning DisorderErika SierraNo ratings yet
- FragileX SyndromeDocument2 pagesFragileX SyndromeAreef MuarifNo ratings yet
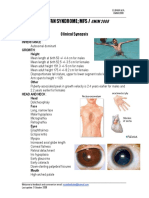
- Marfan SyndromeDocument3 pagesMarfan SyndromeAreef MuarifNo ratings yet
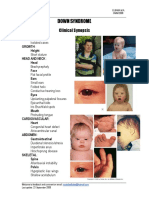
- Down SyndromeDocument2 pagesDown SyndromeAreef MuarifNo ratings yet
- DiGeorge SyndromeDocument3 pagesDiGeorge SyndromeAreef MuarifNo ratings yet
- CHARGE AssociationDocument2 pagesCHARGE AssociationAreef MuarifNo ratings yet
- Angelman Syndrome As Angelman Syndrome As Angelman Syndrome As Angelman Syndrome AsDocument2 pagesAngelman Syndrome As Angelman Syndrome As Angelman Syndrome As Angelman Syndrome AsAreef MuarifNo ratings yet
- Dede2 PretermDocument9 pagesDede2 PretermAreef MuarifNo ratings yet
- Journal 1 PDFDocument9 pagesJournal 1 PDFAntónio Ayrton WidiastaraNo ratings yet
- Pediatric CardiologyDocument6 pagesPediatric CardiologyAreef MuarifNo ratings yet
- Manajemen Hearing Loss PDFDocument15 pagesManajemen Hearing Loss PDFAreef MuarifNo ratings yet
- Neurodevelopment CMV PDFDocument8 pagesNeurodevelopment CMV PDFAreef MuarifNo ratings yet
- gjmbsv3n5 16Document6 pagesgjmbsv3n5 16Areef MuarifNo ratings yet
- Offline - POSBINDU Meri 10Document21 pagesOffline - POSBINDU Meri 10Areef MuarifNo ratings yet
- DikoDocument7 pagesDikoAreef MuarifNo ratings yet
- Dede 5Document5 pagesDede 5Areef MuarifNo ratings yet
- Hiperbilirubinemia by Ucsf HospitalDocument3 pagesHiperbilirubinemia by Ucsf HospitalGalenica HoneyNo ratings yet
- 85120030402Document3 pages85120030402Areef MuarifNo ratings yet
- NICE Guidelines PDFDocument41 pagesNICE Guidelines PDFglruvalNo ratings yet
- Growth PredictionDocument101 pagesGrowth PredictionKristty Magallanes100% (1)
- 1583 - Intermediate Grammar Test 22Document4 pages1583 - Intermediate Grammar Test 22SabinaNo ratings yet
- July 2013 (English)Document28 pagesJuly 2013 (English)Matthew JonesNo ratings yet
- Food and Exercise LogDocument24 pagesFood and Exercise LogmhetfieldNo ratings yet
- Crabtales 061Document20 pagesCrabtales 061Crab TalesNo ratings yet
- Denumire Comerciala DCI Forma Farmaceutica ConcentratieDocument4 pagesDenumire Comerciala DCI Forma Farmaceutica ConcentratieAlina CiugureanuNo ratings yet
- Montelukast SodiumDocument10 pagesMontelukast SodiumikhasilNo ratings yet
- IM Injuction: Definition of Intramuscular InjectionDocument8 pagesIM Injuction: Definition of Intramuscular InjectiondhanasundariNo ratings yet
- Pharyngitis Laryngitis TonsillitisDocument10 pagesPharyngitis Laryngitis Tonsillitisapi-457923289No ratings yet
- CPV/CCV Ag (3 Lines) : VcheckDocument2 pagesCPV/CCV Ag (3 Lines) : VcheckFoamfab WattikaNo ratings yet
- Tacoma-Pierce County Health Department - E-Cigarette E-Mails April 2011 Part 1Document292 pagesTacoma-Pierce County Health Department - E-Cigarette E-Mails April 2011 Part 1American Vaping AssociationNo ratings yet
- TrichDocument30 pagesTrichwadige4668No ratings yet
- Introduction To Dental Anatomy: DR - Yad Raouf BDS, Efb, MrcsedDocument27 pagesIntroduction To Dental Anatomy: DR - Yad Raouf BDS, Efb, MrcsedRabarNo ratings yet
- Ergogenic Aids 1Document24 pagesErgogenic Aids 1unattractiveyouNo ratings yet
- Gallbladder and Bile Duct Anatomy, Function and DiseasesDocument16 pagesGallbladder and Bile Duct Anatomy, Function and DiseasesKadenceFreya-Charisse G PosadasBulintao100% (2)
- R - MN R - 1 Z: Mkg-Xixltechlib/1 T/06Document48 pagesR - MN R - 1 Z: Mkg-Xixltechlib/1 T/06Jhantu MazumderNo ratings yet
- Respiratory Distress SyndromeDocument54 pagesRespiratory Distress SyndromeKristine CaringalNo ratings yet
- Review of drug-induced gingival overgrowthsDocument14 pagesReview of drug-induced gingival overgrowthsRobins DhakalNo ratings yet
- MRI Monitoring System Provides Safety and MobilityDocument4 pagesMRI Monitoring System Provides Safety and MobilityAchiyat WinataNo ratings yet
- Niyog-Niyogan - Quisqualis Indica Herbal Medicine-Health Benefits-Side Effects PDFDocument4 pagesNiyog-Niyogan - Quisqualis Indica Herbal Medicine-Health Benefits-Side Effects PDFJikka RodriguezNo ratings yet
- Referat Arlha PemDocument65 pagesReferat Arlha PemDicky Delonge LesmanaNo ratings yet
- Adverse Reactions To DiureticsDocument8 pagesAdverse Reactions To DiureticsDimas RizkyNo ratings yet
- DHI Score PDFDocument2 pagesDHI Score PDFAnonymous HNGH1oNo ratings yet
- R3 Vital Pulp Therapy With New MaterialsDocument7 pagesR3 Vital Pulp Therapy With New MaterialsWening TyasNo ratings yet